
LINK QUIZZ DO GEMINI: https://gemini.google.com/share/f126f814cea1
Texto na cor preta = Conteúdo dos slides
Vermelho = Explicação por IA
MATERIAL GENÉTICO E SEUS PROCESSAMENTOS
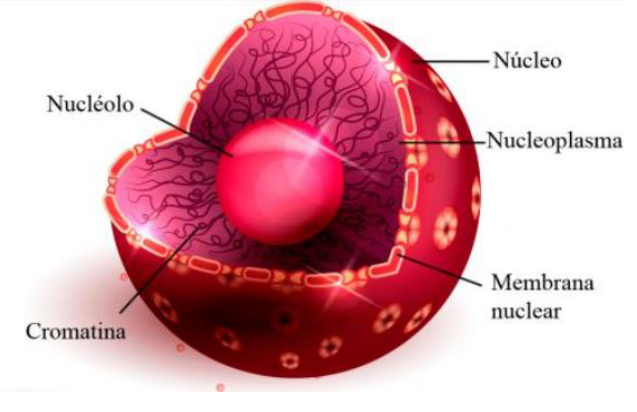
NÚCLEO CELULAR
-
É a região da célula EUCARIONTE onde se encontra o material genético, responsável pela transmissão de características entre gerações.
-
Há também o nucleoplasma, nucléolo e a carioteca.
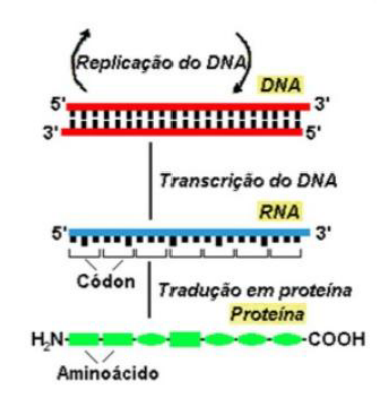
DNA (ÁCIDO DESOXIRRIBONUCLEICO)
-
A estrutura do DNA está associada à sua função: armazenar a informação genética.
-
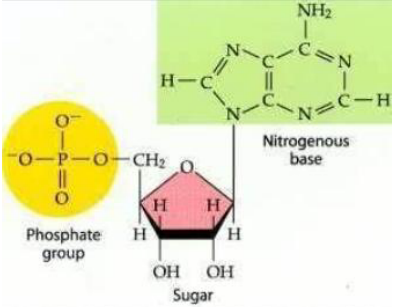
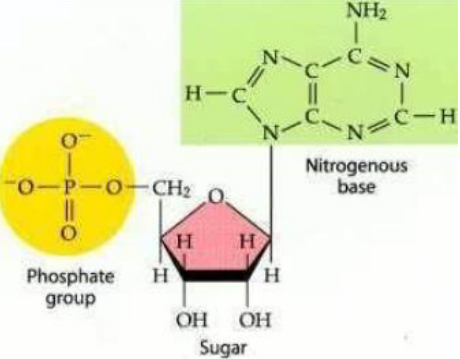
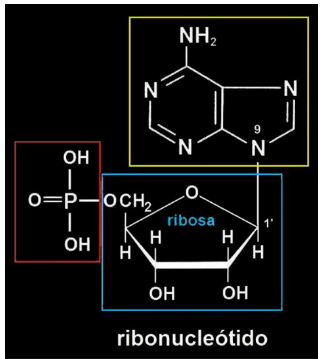
Como um desoxirribonucleotídeo é formado?
-
A pentose e a base nitrogenada são unidas por uma ligação glicosídica = nucleosídeo.
-
Quando o nucleosídeo se liga ao grupamento fosfato, forma um nucleotídeo.
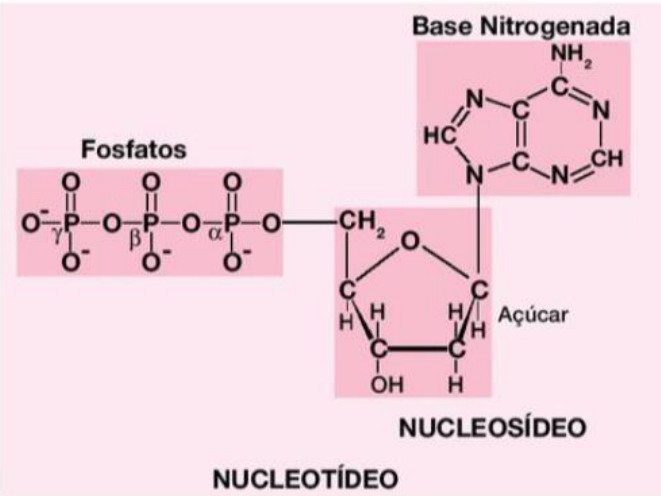
Pergunta de prova: Quais as partes que formam um nucleotídeo:
1. Pentose: É um açúcar. No DNA, essa pentose é a desoxirribose.
2. Base nitrogenada: É uma molécula que faz parte da informação genética.
3. Grupamento fosfato: É outra parte do nucleotídeo, importante para formar a estrutura da molécula.
ÁCIDOS NUCLEICOS
-
O DNA e o RNA são polímeros lineares longos, denominados ácidos nucleicos, que desempenham muitas funções no organismo.
-
Essas macromoléculas consistem em um grande número de nucleotídeos ligados uns aos outros.
-
Cada nucleotídeo é formado por uma pentose, um fosfato e uma base.
Os ácidos nucleicos são moléculas grandes presentes nas células. Os dois principais são o DNA e o RNA. Eles recebem esse nome porque estão relacionados ao armazenamento e ao uso das informações genéticas da célula.
O DNA e o RNA são chamados de polímeros lineares longos porque são formados por várias unidades menores ligadas em sequência, como se fossem uma corrente. Cada “elo” dessa corrente é chamado de nucleotídeo.
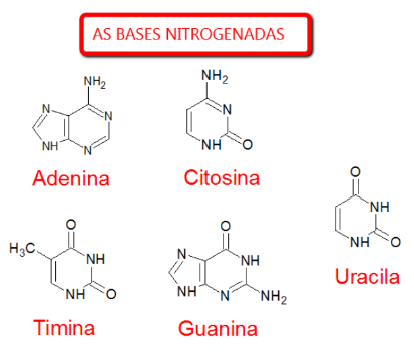
AS BASES NITROGENADAS
-
Adenina, Citosina, Timina, Guanina, Uracila.
-
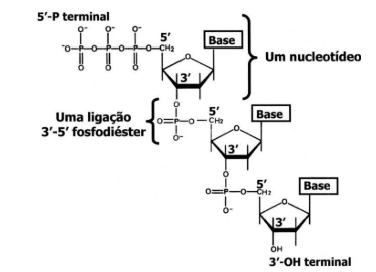
Os desoxirribonucleotídeos estão ligados entre si por pontes fosfodiéster, para formar uma cadeia.
-
Cada ponte fosfodiéster é formada pela extremidade 3′-hidroxila da pentose de um nucleotídeo e a extremidade 5′-fosfato do nucleotídeo adjacente.
-
A cadeia terá no início uma extremidade livre com um grupamento 5′-fosfato (5’P) e na outra extremidade um grupamento 3′-hidroxila (3’OH).
-
Se novos nucleotídeos forem adicionados na cadeia, eles necessariamente precisam se ligar na extremidade 3′.
-
A síntese da cadeia sempre ocorre no sentido 5′ => 3′.
As bases nitrogenadas são partes dos nucleotídeos. No material aparecem cinco: adenina, citosina, timina, guanina e uracila.
De forma bem prática:
- No DNA aparece a timina;
- No RNA aparece a uracila no lugar da timina.
Os desoxirribonucleotídeos são os nucleotídeos que formam o DNA. Eles se ligam uns aos outros por uma ligação chamada ponte fosfodiéster. Essa ligação une um nucleotídeo ao próximo, formando uma cadeia, como se fossem vários elos conectados.
Essa ligação acontece entre duas partes específicas: a extremidade 3’ hidroxila da pentose de um nucleotídeo e a extremidade 5’ fosfato do nucleotídeo seguinte. Em termos simples: uma ponta de um nucleotídeo se encaixa com a ponta do próximo.
Por isso, a cadeia de DNA tem uma direção. Uma ponta é chamada de 5’, porque tem o grupamento fosfato livre. A outra ponta é chamada de 3’, porque tem o grupamento hidroxila livre.
A parte mais importante para prova é: novos nucleotídeos só conseguem ser adicionados na extremidade 3’. Por isso, dizemos que a síntese da cadeia sempre acontece no sentido 5’ → 3’.
IMPORTANTE!
Regra principal
- DNA usa T (timina)
- RNA usa U (uracila)
Ou seja:
- Se é DNA, aparece T
- Se é RNA, aparece U
1. Quando é DNA com DNA
Isso acontece na replicação do DNA.
As combinações são:
- A liga com T
- C liga com G
Então:
- A ↔ T
- C ↔ G
Exemplo:
DNA: A T C G
Fita complementar: T A G C
2. Quando o DNA forma RNA
Isso acontece na transcrição.
Aqui entra o U, porque o RNA não usa T.
As combinações ficam:
- A do DNA liga com U do RNA
- T do DNA liga com A do RNA
- C do DNA liga com G do RNA
- G do DNA liga com C do RNA
Então:
- DNA A → RNA U
- DNA T → RNA A
- DNA C → RNA G
- DNA G → RNA C
Exemplo:
DNA: A T C G
RNA: U A G C
3. Quando é RNA com RNA
Por exemplo, no encaixe entre códon do RNA mensageiro e anticódon do RNA transportador.
Aí o pareamento é:
- A liga com U
- C liga com G
Então:
- A ↔ U
- C ↔ G
Exemplo:
Códon do RNAm: A U G
Anticódon do RNAt: U A C
Esquema-resumo final
Se for DNA:
- A → T
- T → A
- C → G
- G → C
Se estiver formando RNA a partir do DNA:
- A → U
- T → A
- C → G
- G → C
Se for RNA com RNA:
- A → U
- U → A
- C → G
- G → C
ESTRUTURA TRIDIMENSIONAL DO DNA – MODELO PROPOSTO POR WATSON E CRICK (1953)
-
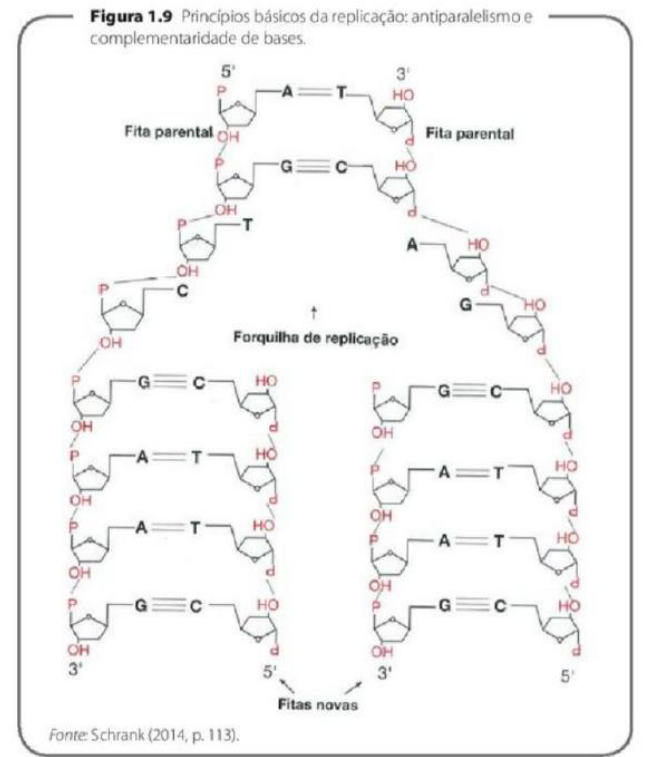
A molécula de DNA apresenta a forma de uma dupla hélice, em que há cadeias antiparalelas enroladas entre si.
-
Ocorre pareamento específico entra as bases nitrogenadas de cada fita – manutenção da hélice.
-
Propriedade de complementaridade.
-
Desnaturação e Renaturação.
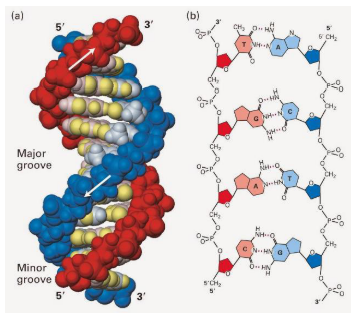
Esse modelo mostra que o DNA tem formato de dupla hélice, parecido com uma escada torcida.
Essa “escada” é formada por duas cadeias de nucleotídeos enroladas uma na outra. Essas cadeias são chamadas de antiparalelas, porque ficam em sentidos opostos: uma vai no sentido 5’ → 3’ e a outra no sentido 3’ → 5’.
As duas fitas se mantêm unidas pelo pareamento específico das bases nitrogenadas. No DNA, a adenina pareia com timina e a citosina pareia com guanina. Então a regra é: A com T e C com G.
A complementaridade significa que uma fita “combina” com a outra. Se uma fita tem a sequência A T C G, a outra obrigatoriamente terá T A G C. Por isso, uma fita pode servir como molde para formar a outra.
A desnaturação acontece quando as duas fitas do DNA se separam, geralmente por alteração de temperatura ou outras condições. Já a renaturação é quando as fitas conseguem se unir novamente, voltando ao pareamento correto entre as bases.
A ideia principal é: o DNA é uma dupla hélice formada por duas fitas antiparalelas e complementares, unidas pelo pareamento das bases A-T e C-G.
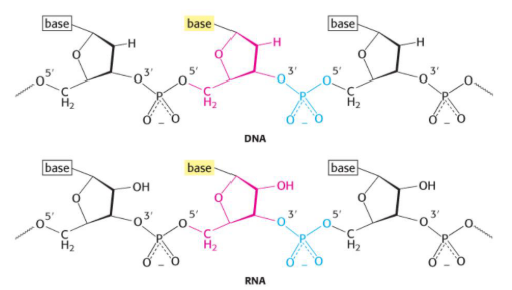
RNA (ÁCIDO RIBONUCLEICO)
-
Também é um polímero formado por nucleotídeos, unidos entre si por ligações fosfodiéster.
-
Possui ribose ao invés de desoxirribose, possui Uracila ao invés de Timina e apresenta uma fita simples e não uma dupla hélice como o DNA.
-
Pareamentos internos – grampos.
-
Regiões em que não há pareamento – alças.
-
Pareamentos – complementaridade.
O RNA, ou ácido ribonucleico, também é um ácido nucleico, assim como o DNA. Ele é formado por vários nucleotídeos ligados uns aos outros por ligações fosfodiéster, formando uma cadeia.
A principal diferença é que o RNA possui ribose como açúcar, enquanto o DNA possui desoxirribose. Além disso, no RNA aparece a base uracila, enquanto no DNA aparece a timina. Então, no RNA, o pareamento acontece com A e U, e não com A e T.
Outra diferença importante é que o RNA geralmente apresenta fita simples, enquanto o DNA tem dupla hélice. Porém, mesmo sendo uma fita simples, o RNA pode se dobrar sobre si mesmo e formar regiões de pareamento interno.
Quando partes da própria molécula de RNA se pareiam, elas formam estruturas chamadas grampos. É como se a fita dobrasse e algumas bases encontrassem suas bases complementares dentro da própria molécula.
As regiões em que não ocorre pareamento recebem o nome de alças. Ou seja, no grampo, uma parte está pareada e outra parte fica “solta”, sem pareamento.
A ideia principal é: o RNA é uma fita simples formada por nucleotídeos, possui ribose e uracila, e pode formar pareamentos internos por complementaridade, criando grampos e alças.
TIPOS DE RNA PRESENTES NA CÉLULA
-
RNA mensageiro (mRNA) – corresponde a 1-5% do RNA total da célula; leva a informação genética do DNA até os ribossomos.
-
RNA ribossômico (rRNA) – corresponde a 75% do RNA total da célula; constitui o principal componente dos ribossomos e é rico em pareamentos internos.
-
RNA transportador (tRNA) – corresponde a 10-15% do RNA total na célula; leva os aminoácidos até os ribossomos para a síntese proteica.
-
Os três participam da síntese proteica.
-
Somente o mRNA é codificador.
Existem três tipos principais de RNA na célula: RNA mensageiro, RNA ribossômico e RNA transportador. Os três participam da síntese proteica, ou seja, do processo de produção de proteínas.
O RNA mensageiro, ou mRNA, representa uma pequena parte do RNA total da célula, cerca de 1 a 5%. Sua função é levar a informação genética que veio do DNA até os ribossomos. Ele funciona como uma “mensagem” com a receita para formar uma proteína.
O RNA ribossômico, ou rRNA, é o mais abundante, representando cerca de 75% do RNA total da célula. Ele forma a maior parte dos ribossomos, que são as estruturas onde ocorre a produção de proteínas. Ele também é rico em pareamentos internos, formando dobramentos na molécula.
O RNA transportador, ou tRNA, representa cerca de 10 a 15% do RNA total da célula. Sua função é levar os aminoácidos até os ribossomos. Esses aminoácidos serão unidos para formar as proteínas.
A parte mais importante é entender que os três RNAs trabalham juntos: o mRNA leva a mensagem, o rRNA forma o ribossomo, e o tRNA traz os aminoácidos.
Quando o material diz que somente o mRNA é codificador, significa que apenas o RNA mensageiro carrega a sequência de informação que será lida para formar a proteína. O rRNA e o tRNA participam do processo, mas não carregam o “código” da proteína.
CROMATINA E CROMOSSOMO
-
Representam a mesma identidade biológica, o ácido nucleico, no entanto, apresentam-se nas células em momentos diferentes e com morfologia e fisiologia próprias.
-
Cromatina (do grego croma, cor) designa a porção do núcleo, com exceção do nucléolo, que se cora e é visível ao microscópio de luz.
-
Ela é um complexo de DNA, proteínas histônicas e não-histônicas, presentes no núcleo em intérfase.
-
Durante a divisão celular, a cromatina sofre alterações em sua morfologia, composição e função, apresentando-se altamente compactada sob a forma de unidades individualizadas conhecidas por cromossomos.
-
Os cromossomos autossômicos geralmente ocorrem aos pares em células somáticas, tanto de animais como de vegetais.
-
O número de cromossomos de uma espécie é constante.
-
A determinação do número de cromossomos de uma espécie geralmente é efetuada na metáfase.
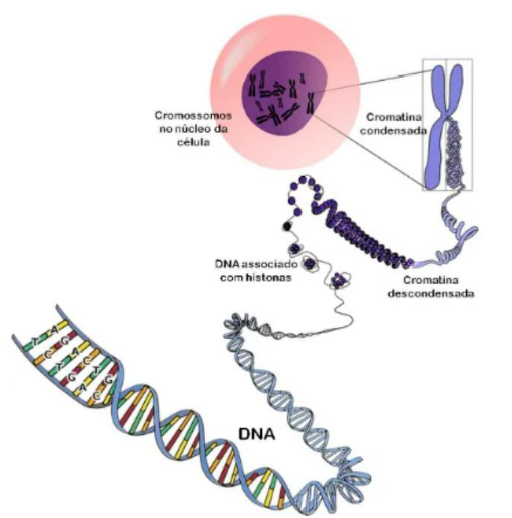
Cromatina e cromossomo são formas diferentes de organização do mesmo material genético: o DNA associado a proteínas. Ou seja, não são coisas totalmente diferentes; o que muda é o momento da célula e o grau de compactação desse material.
A cromatina é a forma mais “desenrolada” e menos compactada do DNA dentro do núcleo. Ela aparece principalmente quando a célula está em intérfase, que é o período em que a célula não está se dividindo. Nessa fase, o DNA precisa estar mais acessível para realizar suas funções.
A cromatina é formada por DNA, proteínas histônicas e proteínas não-histônicas. As histonas ajudam a organizar e compactar o DNA, como se servissem de suporte para enrolar esse material.
Durante a divisão celular, a cromatina se compacta bastante. Quando fica altamente condensada, passa a ser vista como cromossomo. Então, o cromossomo é a cromatina em uma forma mais organizada, compacta e individualizada.
Uma forma simples de pensar é: cromatina é o DNA mais solto; cromossomo é o DNA bem compactado para a divisão celular.
Os cromossomos autossômicos são aqueles que geralmente aparecem aos pares nas células somáticas, ou seja, nas células do corpo. Isso ocorre tanto em animais quanto em vegetais.
Cada espécie possui um número constante de cromossomos. Por exemplo, uma espécie tem sempre um número característico de cromossomos, que não muda de indivíduo para indivíduo daquela espécie em condições normais.
A contagem dos cromossomos geralmente é feita na metáfase, porque nessa fase da divisão celular eles estão bem compactados e visíveis ao microscópio.
A ideia principal é: cromatina e cromossomo são o mesmo material genético em estados diferentes: na intérfase aparece como cromatina; na divisão celular aparece compactado como cromossomo.
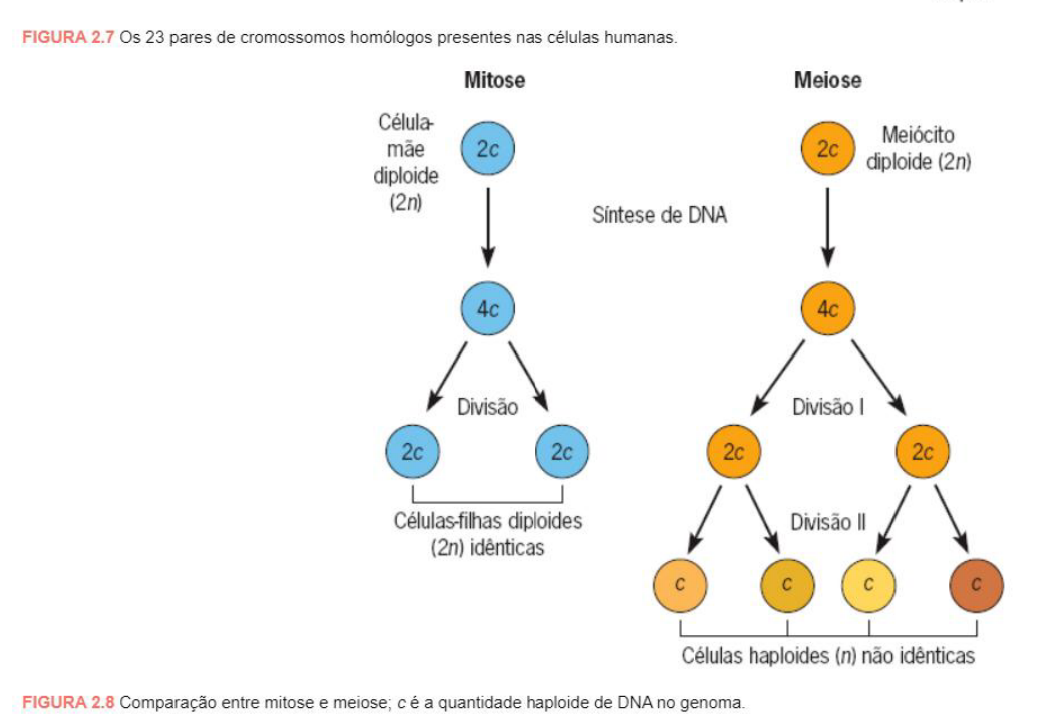
HUMANOS
-
23 pares de cromossomos, formados por DNA.
-
22 pares são autossômicos.
-
1 par é formado pelos 2 cromossomos sexuais.
-
Em cada cromossomo há uma molécula de DNA de dupla fita associada firmemente a histonas.
-
Histonas – empacotamento do DNA e na expressão gênica.
-
Intérfase – cromatina relativamente descondensada e dispersa pelo núcleo (exceção: heterocromatina – 10% da cromatina na intérfase – centrômeros, telômeros).
-
Mitose – cromatina extremamente condensada para que os cromossomos sejam distribuídos corretamente para as células-filha.

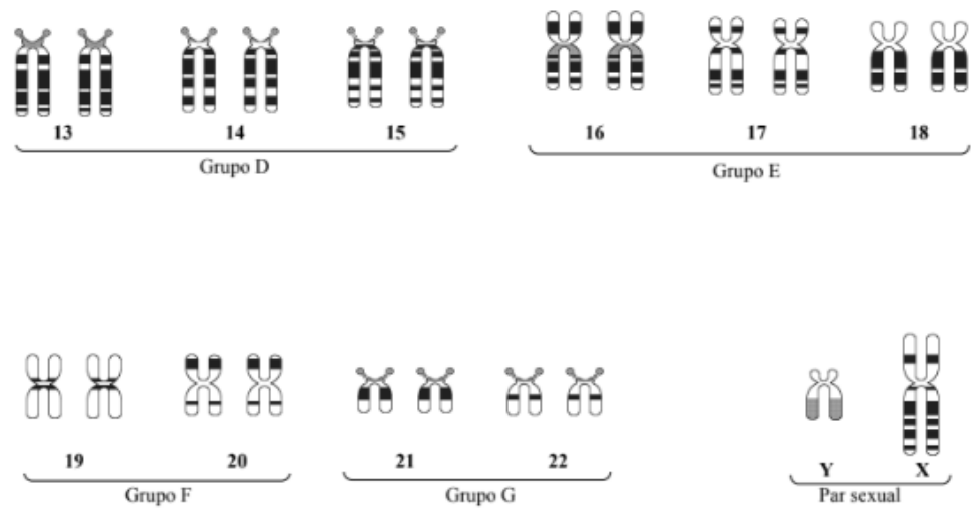
Nos seres humanos, as células somáticas, ou seja, as células do corpo, possuem 23 pares de cromossomos, totalizando 46 cromossomos. Esses cromossomos são formados por DNA associado a proteínas.
Desses 23 pares, 22 pares são autossômicos. Isso significa que eles não determinam diretamente o sexo biológico. Eles carregam informações relacionadas a várias características do organismo.
O último par é formado pelos cromossomos sexuais. Esse par está relacionado à determinação sexual. Portanto, o ser humano possui 22 pares autossômicos + 1 par sexual.
Cada cromossomo contém uma molécula de DNA de dupla fita. Essa molécula fica associada a proteínas chamadas histonas, que ajudam a organizar e compactar o DNA dentro do núcleo.
As histonas são importantes para o empacotamento do DNA. Como o DNA é muito longo, ele precisa ficar bem organizado dentro do núcleo. Além disso, as histonas também participam da expressão gênica, ou seja, ajudam a controlar quais partes do DNA serão mais ou menos utilizadas pela célula.
Na intérfase, a cromatina fica relativamente descondensada e espalhada pelo núcleo. Isso facilita o uso das informações genéticas pela célula. Porém, uma pequena parte permanece mais compactada, chamada de heterocromatina.
A heterocromatina representa cerca de 10% da cromatina na intérfase e está presente em regiões como centrômeros e telômeros. De forma simples, são regiões do cromossomo que permanecem mais compactadas mesmo quando a célula não está se dividindo.
Durante a mitose, a cromatina fica extremamente condensada, formando cromossomos bem visíveis. Isso é necessário para que o material genético seja separado corretamente e distribuído para as células-filhas.
A ideia principal é: na intérfase, o DNA fica mais solto como cromatina; na mitose, ele se compacta em cromossomos para ser dividido corretamente entre as novas células.
REPLICAÇÃO DO DNA
-
Processo essencial para a duplicação do material genético, que ocorre durante a divisão celular em todos os organismos.
-
Permite que o mesmo material genético do organismo original esteja presente nas duas células filhas.
-
Fita molde.
-
Fragmento deve se parear com a fita molde e ter uma extremidade 3′-OH livre.
REPLICAÇÃO DO DNA
-
4 CARACTERÍSTICAS IMPORTANTES: É semidescontínua. É semiconservativa. Ocorre sempre no sentido 5′-3′. Sempre inicia em sequências específicas (origens).
-
A dupla hélice de DNA deverá ser aberta em um trecho, formando a chamada Bolha de replicação.
-
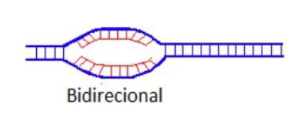
A partir disso, a replicação pode ser uni ou bidirecional.
-
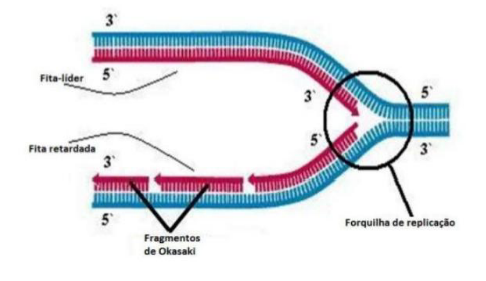
Replicação unidirecional – um conjunto de proteínas chamada replissomo começa na origem que é denominada forquilha de replicação.
-
Replicação bidirecional – Existem 2 forquilhas de replicação, que partem da origem e seguem em direções opostas.
-
Princípios básicos da replicação: antiparalelismo e complementaridade de bases.
-
Em cada uma das fitas será produzida uma fita nova complementar, seguindo o pareamento AT e CG, o sentido 5′-3′ e a conformação antiparalela em relação à fita molde.
A replicação do DNA tem quatro características importantes: é semidescontínua, semiconservativa, ocorre sempre no sentido 5’ → 3’ e começa em regiões específicas chamadas origens de replicação.
COMPONENTES DA REPLICAÇÃO
-
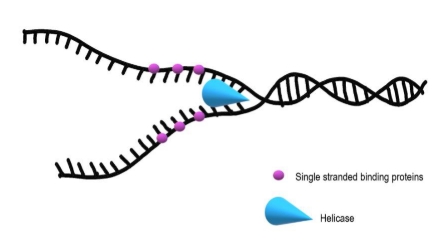
Helicases – realiza a abertura da dupla hélice, permitindo a movimentação da forquilha de replicação.
-
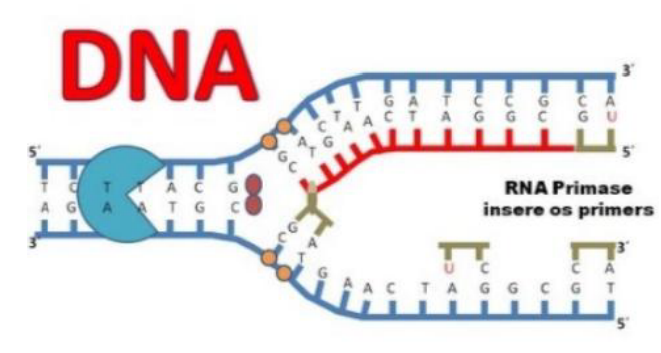
Primers (iniciadores) – seguimentos de ácido nucleico que se associa a fita parental e permite a adição de nucleotídeos a partir dele (possui uma extremidade 3′-OH livre).
-
DNA polimerases – realizam a incorporação dos nucleotídeos na extremidade 3′-OH livre.
-
SSB (Single-stranded DNA biding protein) – mantem as fitas de DNA separadas.
-
Topoisomerase – Enzimas responsáveis por reduzir a tensão gerada pelas supertorções, induzindo uma quebra transitória nessa cadeia.
Os componentes da replicação são as moléculas e enzimas que ajudam o DNA a ser copiado corretamente. Cada uma tem uma função específica no processo.
A helicase é a enzima que abre a dupla hélice do DNA. Ela separa as duas fitas, permitindo que cada uma sirva como molde para a formação de uma nova fita. Essa abertura forma a região chamada forquilha de replicação.
Os primers, ou iniciadores, são pequenos segmentos de ácido nucleico que se ligam à fita parental. Eles são importantes porque fornecem uma extremidade 3’-OH livre, que é o ponto de partida para a adição dos novos nucleotídeos.
As DNA polimerases são as enzimas que adicionam os nucleotídeos novos à cadeia em formação. Elas só conseguem fazer isso a partir da extremidade 3’-OH livre, por isso a presença do primer é necessária.
As proteínas SSB se ligam às fitas simples de DNA depois que elas são separadas. A função delas é manter essas fitas abertas, impedindo que elas se unam novamente antes da replicação acontecer.
A topoisomerase atua para aliviar a tensão causada pela abertura da dupla hélice. Quando o DNA vai sendo desenrolado, surgem torções na molécula; a topoisomerase reduz essa tensão fazendo quebras temporárias na cadeia.
A ideia principal é: a helicase abre o DNA, as SSB mantêm as fitas separadas, a topoisomerase alivia a tensão, o primer dá o ponto de partida e a DNA polimerase adiciona os novos nucleotídeos.
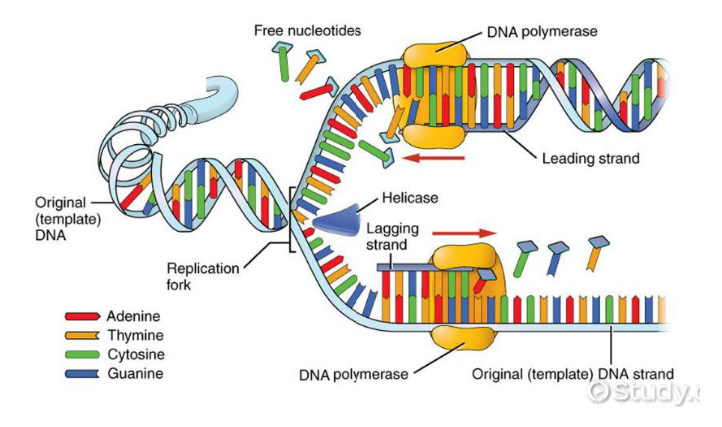
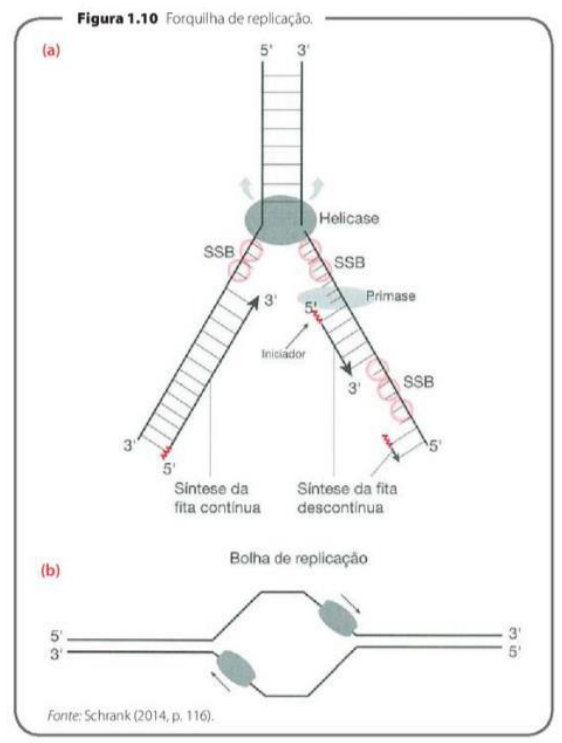
A REPLICAÇÃO É UM PROCESSO SEMIDESCONTÍNUO
-
A DNA-polimerase realiza a síntese somente no sentido 5′-3′.
-
Uma das fitas novas é sintetizada de forma contínua a partir da fita molde que está na posição 3′-5′.
-
A outra fita é sintetizada de forma descontínua, uma vez que a fita-molde está na posição 5′-3′.
-
Fita contínua – necessário apenas um iniciador.
-
Fita descontínua – são necessários vários iniciadores, cada um contribuindo para a síntese de um fragmento, chamado de fragmento de Okasaki.
-
A DNA-ligase une esses fragmentos de Okasaki da fita descontínua.
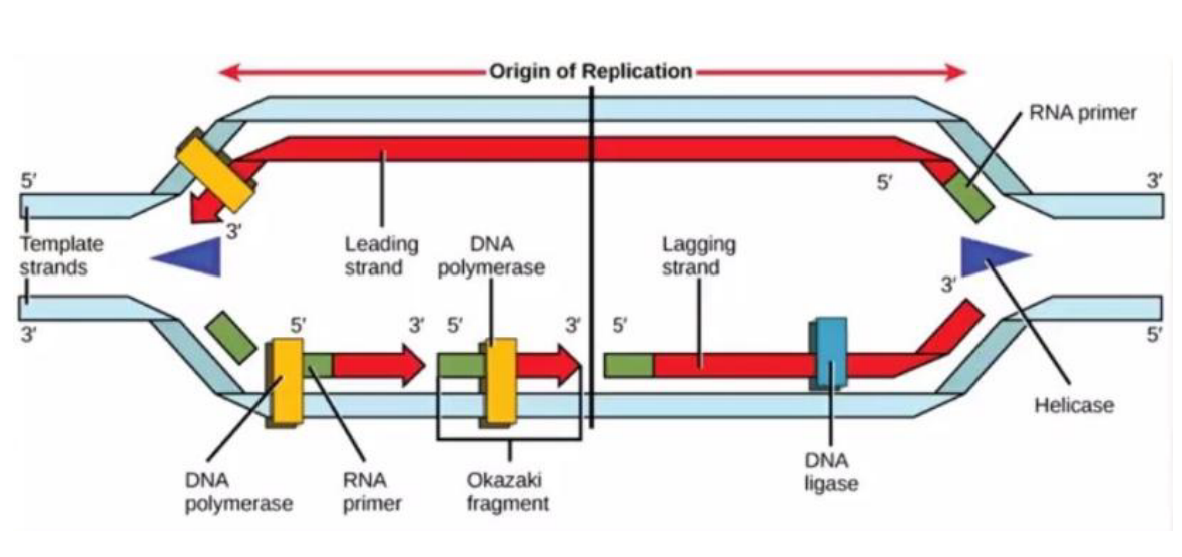
A replicação é semidescontínua porque as duas fitas novas do DNA não são produzidas do mesmo jeito. Isso acontece porque a DNA-polimerase só consegue sintetizar DNA no sentido 5’ → 3’.
Como as fitas do DNA são antiparalelas, uma fita molde está no sentido 3’ → 5’ e a outra está no sentido 5’ → 3’. Isso interfere diretamente na forma como a nova fita será montada.
Na fita molde que está no sentido 3’ → 5’, a nova fita consegue ser produzida de maneira contínua, seguindo normalmente o sentido 5’ → 3’. Por isso, essa é chamada de fita contínua. Ela precisa de apenas um iniciador.
Já na outra fita molde, que está no sentido 5’ → 3’, a produção não consegue acontecer de forma direta e contínua. Por isso, a nova fita é feita em pequenos pedaços. Essa é chamada de fita descontínua.
Na fita descontínua, são necessários vários iniciadores, porque cada pedaço precisa de um ponto de partida. Cada pedaço formado recebe o nome de fragmento de Okazaki.
Depois que esses fragmentos são produzidos, entra a DNA-ligase, que funciona como uma “cola”. Ela une os fragmentos de Okazaki, formando uma fita contínua ao final do processo.
A ideia principal é: uma fita nova é feita direto, de forma contínua; a outra é feita em pedaços, chamados fragmentos de Okazaki, que depois são unidos pela DNA-ligase.

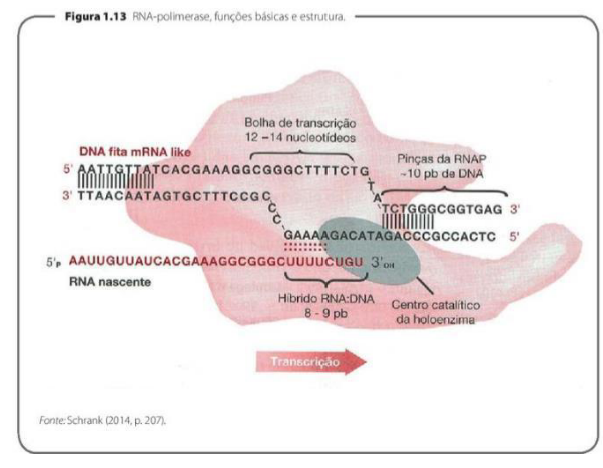
TRANSCRIÇÃO EM EUCARIONTES
-
É o processo pelo qual uma molécula de RNA é sintetizada a partir de uma das fitas da molécula de DNA.
-
As bases ACTG do DNA são pareadas, respectivamente, com as bases UGAC do RNA.
-
Os nucleotídeos da fita de RNA são unidos uns aos outros por ligação fosfodiéster e depende da ação das RNA polimerases.
A transcrição em eucariontes é o processo em que a célula usa uma das fitas do DNA como molde para produzir uma molécula de RNA. Ou seja, a informação que estava no DNA é “copiada” em forma de RNA.
Durante esse processo, as bases do DNA se pareiam com bases complementares do RNA. A diferença principal é que, no RNA, não existe timina, entra uracila no lugar dela.
Então, o pareamento fica assim:
DNA A → RNA U
DNA C → RNA G
DNA T → RNA A
DNA G → RNA C
Por isso o material diz que as bases A C T G do DNA são pareadas, respectivamente, com U G A C do RNA.
Os nucleotídeos que vão formando a fita de RNA são ligados entre si por ligações fosfodiéster, do mesmo tipo geral que aparece na formação das cadeias de DNA e RNA.
Esse processo depende das RNA polimerases, que são as enzimas responsáveis por montar a nova fita de RNA a partir da fita molde de DNA.
A ideia principal é: na transcrição, uma fita do DNA serve de molde para formar uma fita de RNA, usando U no lugar de T.
INÍCIO DA TRANSCRIÇÃO E FORMAÇÃO DO COMPLEXO DE PRÉ-INICIAÇÃO
-
A RNA polimerase identifica o local onde ela deve iniciar a transcrição por uma região específica do DNA, chamada de promotor.
-
Sequência chamada TATA box está localizada anteriormente aos genes e é reconhecida pela RNA polimerase.
-
Forma-se então uma estrutura entre o DNA e a RNA polimerase, denominada complexo de pré-inciação.
-
A partir deste complexo, é sintetizada a primeira ligação fosfodiéster da molécula de RNA.
O início da transcrição acontece quando a RNA polimerase encontra o ponto certo do DNA onde deve começar a produzir o RNA. Esse ponto não é aleatório: ele fica em uma região específica chamada promotor. O promotor funciona como uma “placa de início” para a RNA polimerase. Ele indica onde a enzima deve se ligar para começar a transcrição de determinado gene.
Dentro dessa região promotora, o material cita a TATA box. Ela é uma sequência específica do DNA que fica antes dos genes e ajuda no reconhecimento do local de início da transcrição.
Quando a RNA polimerase se associa ao DNA nessa região, forma-se o complexo de pré-iniciação. Esse complexo é como uma preparação inicial: a enzima está posicionada no local correto e pronta para começar a formar o RNA.
Depois que esse complexo está formado, a RNA polimerase começa a unir os primeiros nucleotídeos do RNA. A primeira união entre esses nucleotídeos ocorre por uma ligação fosfodiéster.
A ideia principal é: a RNA polimerase reconhece o promotor, se posiciona com ajuda da região TATA box, forma o complexo de pré-iniciação e inicia a síntese do RNA.
ALONGAMENTO DA MOLÉCULA DE RNA
-
A RNA polimerase se desloca ao longo da molécula de DNA com duas ações, desenrolar a dupla hélice e adicionar sequencialmente os nucleotídeos na molécula de RNA que está sendo sintetizada.
-
A RNA polimerase identifica o local onde deve parar a síntese de RNA a partir de sequências específicas da molécula de DNA denominadas finalizadores.
-
A transcrição termina com a dissociação da RNA polimerase do DNA e a liberação da nova molécula de RNA transcrita.
O alongamento da molécula de RNA é a fase em que a RNA polimerase já iniciou a transcrição e agora vai avançando pela molécula de DNA para montar a fita de RNA.
Enquanto se desloca pelo DNA, a RNA polimerase faz duas coisas principais: ela desenrola a dupla hélice em uma pequena região e vai adicionando nucleotídeos um por um na molécula de RNA que está sendo formada.
Esses nucleotídeos são adicionados seguindo a regra de complementaridade: quando a fita molde do DNA tem A, entra U no RNA; quando tem T, entra A; quando tem C, entra G; e quando tem G, entra C.
A RNA polimerase continua esse processo até encontrar uma sequência específica no DNA chamada finalizador. Essa sequência funciona como um sinal de parada, indicando que a síntese daquele RNA deve terminar.
No final da transcrição, a RNA polimerase se solta do DNA e a nova molécula de RNA transcrita é liberada.
A ideia principal é: no alongamento, a RNA polimerase anda pelo DNA, abre a hélice e monta o RNA; quando encontra o finalizador, ela para e libera o RNA formado.
PROCESSAMENTO DO RNA MENSAGEIRO
-
O processo de transcrição produz a molécula de mRNA, a qual precisa ser processada antes da tradução.
-
O produto inicial da transcrição (pré-mRNAs) precisa ser extensamente modificado antes de ser exportado do núcleo.
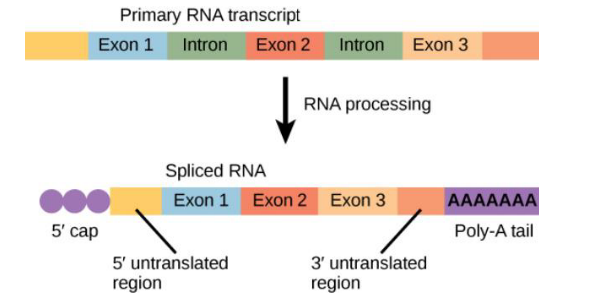
O processamento do RNA mensageiro acontece depois da transcrição. A transcrição produz uma molécula inicial de RNA, chamada de pré-mRNA, mas ela ainda não está pronta para ser usada na tradução.
Antes de sair do núcleo, esse pré-mRNA precisa ser modificado. Essas modificações transformam o pré-mRNA em um mRNA maduro, que poderá ser levado para fora do núcleo e usado na produção de proteína.
Então, a ideia é que a célula primeiro faz uma “versão inicial” da mensagem e depois precisa ajustá-la antes de enviá-la para os ribossomos.
A ideia principal é: a transcrição forma o pré-mRNA; o processamento modifica esse pré-mRNA; depois disso ele pode sair do núcleo e participar da tradução.
ETAPAS ENVOLVIDAS NO PROCESSAMENTO DO RNA MENSAGEIRO
-
Extremidade 5′ do mRNA se agrega a um nucleotídeo metilado chamado cap – o chamado cap é um nucleotídeo específico (7-metilguanosina) que se liga na extremidade 5′ do mRNA – previne que fosfatases e nuclease degradem a extremidade 5′ do mRNA, além de ser necessário para unir o mRNA ao ribossomo no processo da tradução.
-
Extremidade 3′ do mRNA se poliadenina – uma sequência chamada cauda de poli A, de aproximadamente 220 nucleotídeos é adicionada à extremidade 3′ do mRNA – proteção dessa extremidade contra degradação enzimática.
-
Retirada dos íntrons (splicing) e ligação do RNA – a modificação mais marcante dos pré-mRNA é a remoção dos íntrons por splicing.
-
O mRNA é quebrado entre os íntrons (sequências não codificantes) e os éxons (sequências codificantes), em seguida os íntrons são removidos e os éxons são unidos entre si – Splicing.
-
Após o processamento do mRNA ele está preparado para sair do núcleo celular para o citoplasma e seguir o processo de tradução da informação contida no DNA.
O processamento do RNA mensageiro envolve modificações feitas no pré-mRNA antes que ele saia do núcleo. Essas modificações protegem a molécula e deixam o mRNA pronto para participar da tradução.
Na extremidade 5’ do mRNA é adicionado o cap, formado por um nucleotídeo específico chamado 7-metilguanosina. Esse cap funciona como uma proteção contra a degradação da ponta 5’ e também ajuda o mRNA a se ligar ao ribossomo durante a tradução.
Na extremidade 3’ é adicionada a cauda poli-A, formada por uma sequência de vários nucleotídeos de adenina. Essa cauda também tem função de proteção, evitando que a extremidade 3’ seja degradada por enzimas.
Outra etapa importante é o splicing, que é a retirada dos íntrons. Os íntrons são sequências não codificantes, ou seja, partes que não serão usadas diretamente para formar a proteína.
Depois da retirada dos íntrons, os éxons são unidos. Os éxons são as sequências codificantes, ou seja, as partes que permanecem no mRNA maduro e carregam a informação que será usada na tradução.
Então, o processamento do mRNA tem três ideias principais: colocar o cap na ponta 5’, adicionar a cauda poli-A na ponta 3’ e remover os íntrons, unindo os éxons.
A ideia final é: o pré-mRNA precisa ser protegido e editado dentro do núcleo; depois disso, vira mRNA maduro, sai para o citoplasma e segue para a tradução.
TRADUÇÃO
-
Cada mRNA sai para o citoplasma e se combina com um ribossomo, para que tenha início a tradução.
-
Ribossomos – sítios da síntese proteica e usualmente designados de acordo com sua taxa de sedimentação (80S – eucariontes).
-
Esses ribossomos são formados por duas subunidades, uma menor chamada 40S e outra maior chamada 60S (Cada uma constituída por uma ou mais moléculas de rRNA e determinadas proteínas).
-
Na subunidade menor há dois sítios denominados P e A, para a ligação dos tRNA.
A tradução é o processo em que a informação do mRNA é usada para produzir uma proteína. Depois que o mRNA é processado, ele sai do núcleo e vai para o citoplasma, onde se liga a um ribossomo.
Os ribossomos são os locais onde acontece a síntese proteica, ou seja, onde as proteínas são montadas. Nos eucariontes, o ribossomo é chamado de 80S, que é uma forma de classificação relacionada à sua taxa de sedimentação.
Esse ribossomo 80S é formado por duas partes: uma subunidade menor, chamada 40S, e uma subunidade maior, chamada 60S. Cada uma dessas subunidades é formada por rRNA e proteínas.
O rRNA é importante porque faz parte da estrutura do ribossomo. Então, o ribossomo não é feito só de proteína; ele também contém RNA ribossômico.
Na subunidade menor existem dois locais principais de ligação para o tRNA, chamados de sítios P e A. O tRNA é quem traz os aminoácidos que serão usados para formar a proteína.
De forma simples, o mRNA traz a mensagem, o ribossomo lê essa mensagem, e o tRNA traz os aminoácidos para montar a proteína.
A ideia principal é: na tradução, o mRNA se liga ao ribossomo no citoplasma, e os tRNAs ajudam a trazer os aminoácidos para formar uma proteína.
A IMPORTÂNCIA DO RNA TRANSPORTADOR NA SÍNTESE PROTEICA
-
Os tRNA tem a função de captar aminoácidos dispersos na célula e transportá-los até os ribossomos.
-
Código genético – Sequência de 3 nucleotídeos do mRNA constitui um códon.
-
O tRNA – 2 estruturas importantes – aceptor, que se liga ao aminoácido e anticódon, que possui uma sequência de 3 nucleotídeos e é COMPLEMENTAR à sequência do códon.
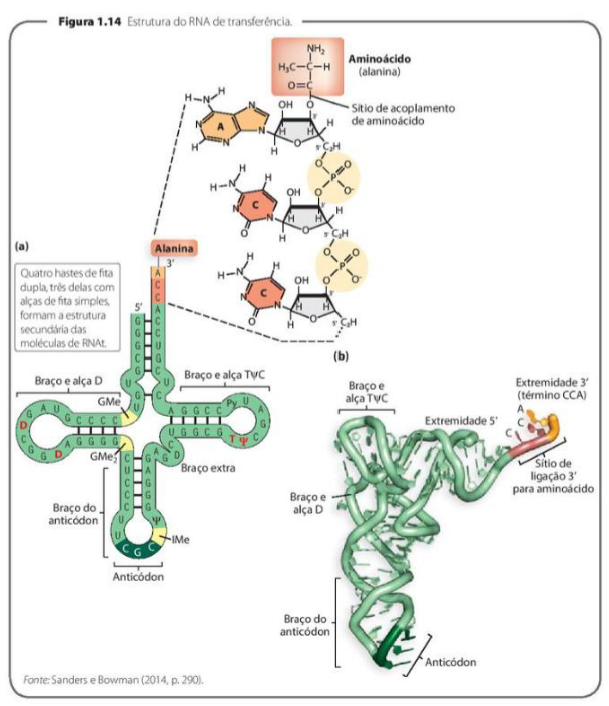
O RNA transportador, ou tRNA, é essencial na síntese proteica porque ele leva os aminoácidos até os ribossomos. Esses aminoácidos ficam dispersos na célula e precisam ser transportados para o local onde a proteína será montada.
O código genético funciona pela leitura do mRNA em grupos de três nucleotídeos. Cada grupo de três bases do mRNA é chamado de códon. Cada códon está relacionado a um aminoácido específico.
O tRNA tem duas regiões muito importantes. A primeira é o aceptor, que é a parte onde o aminoácido se liga. É como se fosse o local onde o tRNA “carrega” o aminoácido.
A segunda região é o anticódon, que possui três nucleotídeos. Esse anticódon é complementar ao códon do mRNA. Ou seja, ele se encaixa no códon seguindo a regra das bases: A com U e C com G.
Por exemplo, se o códon do mRNA for AUG, o anticódon do tRNA será UAC. Assim, o tRNA certo se encaixa no códon certo e leva o aminoácido correto para formar a proteína.
A ideia principal é: o mRNA traz o código em códons, e o tRNA reconhece esses códons pelo anticódon, levando o aminoácido correspondente até o ribossomo.
CÓDON DE INICIAÇÃO
-
A primeira tarefa dos ribossomos é localizar o códon de iniciação, que é o primeiro códon do mRNA a ser traduzido.
-
Esse códon sempre apresenta a sequência AUG, que codifica o aminoácido metionina.
-
A tradução também segue o sentido 5′-3′ da molécula do mRNA.
-
O tRNA correspondente ao códon de iniciação se encaixa no sítio P do ribossomo.
-
Os aminoácidos inseridos serão unidos através de ligações peptídicas.
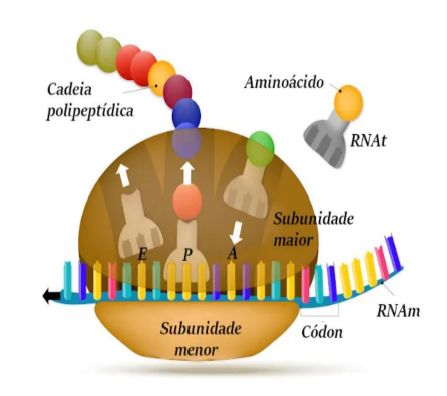
O códon de iniciação é o ponto onde a tradução começa. A primeira tarefa do ribossomo é encontrar, no mRNA, qual é o primeiro códon que deve ser lido.
Esse códon de início é sempre AUG. Ele codifica o aminoácido metionina, que será o primeiro aminoácido colocado na cadeia proteica.
A tradução acontece no sentido 5’ → 3’ do mRNA. Isso significa que o ribossomo lê a molécula de mRNA nessa direção, avançando de códon em códon.
O tRNA correspondente ao AUG possui o anticódon complementar e se encaixa no sítio P do ribossomo. A partir daí, outros tRNAs vão chegando com novos aminoácidos.
Conforme os aminoácidos são trazidos pelos tRNAs, eles são unidos por ligações peptídicas. Essas ligações formam a cadeia de aminoácidos que dará origem à proteína.
A ideia principal é: a tradução começa no códon AUG, que chama o tRNA com metionina para o sítio P; depois os aminoácidos vão sendo unidos por ligações peptídicas.
ALONGAMENTO DA CADEIA PEPTÍDICA
-
A etapa de alongamento é mediada por moléculas denominadas fatores de alongamento e consome energia.
-
O alongamento começa quando o tRNA se encaixa no sítio A do ribossomo.
-
O aminoácido desse tRNA se ligará à metionina que está no sítio P através de uma ligação peptídica.
-
Translocação do mRNA no interior do ribossomo, avançando 3 nucleotídeos por vez.
-
Translocação – o primeiro tRNA se retira do sítio P e vai para o sítio E onde será liberado, o segundo tRNA se desloca do sítio A para o P, um terceiro tRNA chega ao sítio A, dando continuidade ao alongamento.
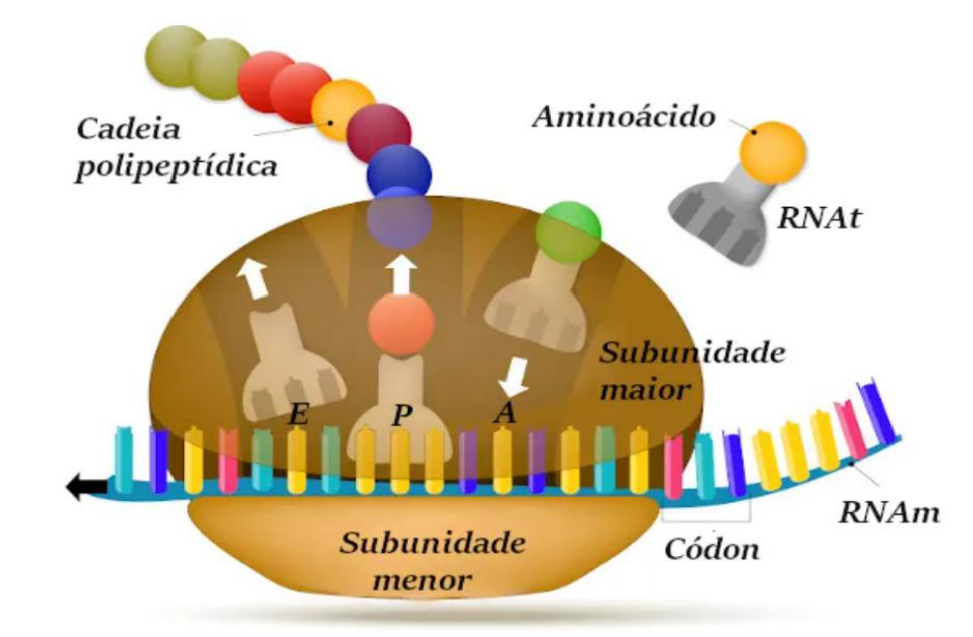
O alongamento da cadeia peptídica é a fase em que a proteína começa a crescer. Depois que a tradução inicia com a metionina no sítio P, novos aminoácidos vão sendo adicionados um por vez.
Essa etapa depende dos fatores de alongamento e consome energia. Isso acontece porque o ribossomo precisa movimentar o mRNA, encaixar novos tRNAs e formar as ligações entre os aminoácidos.
O alongamento começa quando um novo tRNA se encaixa no sítio A do ribossomo. Esse tRNA traz um aminoácido que será unido à metionina, que já está no sítio P.
A união entre os aminoácidos ocorre por uma ligação peptídica. Essa ligação é o que forma a cadeia peptídica, ou seja, a futura proteína.
Depois disso acontece a translocação. O ribossomo avança no mRNA de 3 nucleotídeos por vez, porque cada códon tem três bases.
Durante a translocação, o primeiro tRNA sai do sítio P e vai para o sítio E, onde será liberado. O segundo tRNA, que estava no sítio A, passa para o sítio P. Então um terceiro tRNA chega ao sítio A, trazendo outro aminoácido.
Esse ciclo se repete várias vezes: entra tRNA no A, forma ligação peptídica, o ribossomo anda 3 bases, o tRNA sai pelo E, e a cadeia cresce.
A ideia principal é: no alongamento, o ribossomo lê o mRNA de três em três bases, recebe tRNAs no sítio A, une aminoácidos por ligações peptídicas e vai aumentando a cadeia proteica.
CÓDON DE TERMINAÇÃO
-
O alongamento continua até que o ribossomo atinja o códon de terminação do mRNA (UAA, UGA ou UAG).
-
Quando isso ocorre a síntese da proteína é finalizada.
-
Assim, o sítio ao invés de receber um novo tRNA, é ocupado por um fator de terminação.
-
O polipeptídeo que estava sendo formado se separa do último tRNA, desprendendo-se do mRNA e do ribossomo – proteína é liberada.
-
Subunidades do ribossomo se separam desse mRNA e vão se ligar em outro mRNA para sintetizar uma nova proteína.
O códon de terminação é o sinal que indica que a tradução deve parar. Ele aparece no mRNA quando a proteína já está chegando ao fim de sua formação.
Existem três códons de terminação: UAA, UGA e UAG. Eles não chamam um tRNA com aminoácido. Em vez disso, quando o ribossomo encontra um desses códons, entra um fator de terminação.
Esse fator de terminação ocupa o lugar onde entraria um novo tRNA e faz com que a síntese da proteína seja finalizada. Assim, não são adicionados mais aminoácidos à cadeia.
Depois disso, o polipeptídeo que estava sendo formado se separa do último tRNA. Esse polipeptídeo se desprende do mRNA e do ribossomo, sendo liberado como proteína.
Por fim, as subunidades do ribossomo se separam daquele mRNA. Depois, elas podem se ligar a outro mRNA para iniciar a síntese de uma nova proteína.
A ideia principal é: a tradução começa no AUG e termina quando o ribossomo encontra UAA, UGA ou UAG; nesse momento a proteína é liberada e o ribossomo se desmonta para ser reutilizado.
CÓDIGO GENÉTICO
-
RNA – 4 tipos de nucleotídeos (A, U, C, G).
-
Na célula existem 20 tipos de aminoácidos que podem compor uma proteína.
-
A célula utiliza uma combinação de 3 nucleotídeos (trinca), chamada de códon, para codificar cada aminoácido.
-
O número possível de trincas formados por esses 4 nucleotídeos é de 64.
-
O conjunto desses 64 códons é chamado de código genético.
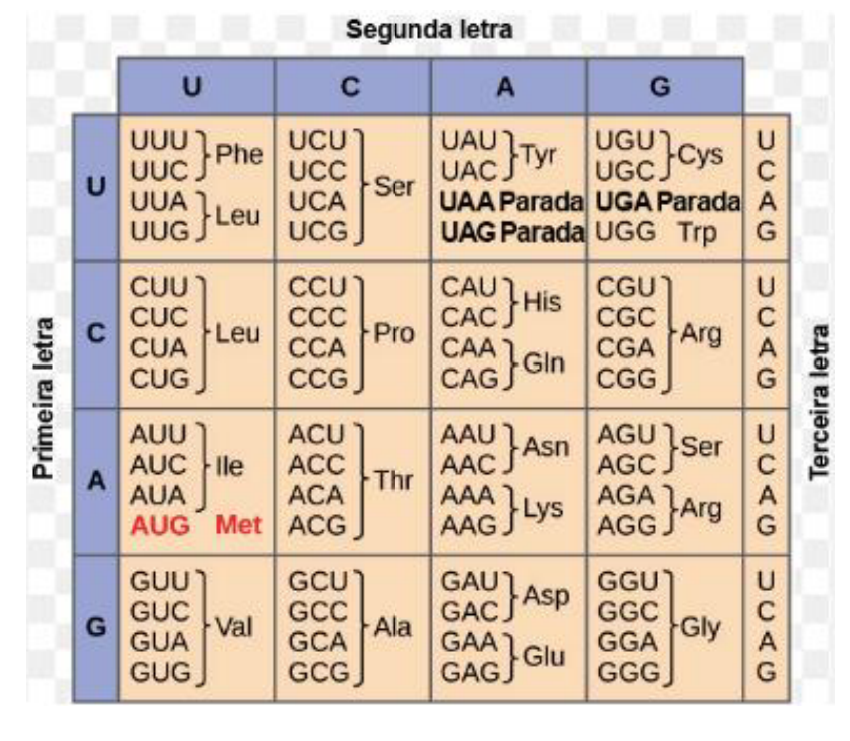
-
3 códons não codificam aminoácido, pois são códons de terminação.
-
61 códons para codificar 20 aminoácidos – a maioria dos aminoácidos (exceto a metionina e o triptofano) pode ser codificada por mais de um códon.
-
Código genético é redundante.
-
Controvérsia se o código genético é ou não universal.
-
Até tempos atrás, acreditava-se que era o mesmo para todos os seres vivos.
-
No entanto, novas pesquisas têm demonstrado que os genes nucleares e mitocondriais em humanos e em outros organismos podem seguir códigos ligeiramente diferentes.
O código genético é o conjunto de regras que a célula usa para transformar a informação do mRNA em uma sequência de aminoácidos, formando uma proteína.
O RNA possui quatro tipos de bases: A, U, C e G. Só que existem 20 tipos de aminoácidos que podem formar proteínas. Então, para conseguir representar todos esses aminoácidos, a célula não lê as bases uma por uma, mas em grupos de três nucleotídeos.
Cada grupo de três bases do mRNA é chamado de códon. Por exemplo, AUG é um códon. Cada códon pode indicar um aminoácido ou pode indicar o fim da tradução.
Como existem 4 bases possíveis e elas são lidas em trincas, o número total de combinações possíveis é 64 códons. Esse conjunto de 64 possibilidades é chamado de código genético.
Desses 64 códons, 3 são códons de terminação: UAA, UGA e UAG. Eles não codificam aminoácidos; servem como sinal para parar a tradução.
Sobram então 61 códons para codificar os 20 aminoácidos. Por isso, vários aminoácidos podem ser representados por mais de um códon. Essa característica é chamada de redundância do código genético.
Quando o material diz que o código genético é redundante, significa que existem “códigos diferentes” que podem levar ao mesmo aminoácido. A exceção citada são a metionina e o triptofano, que são codificados por apenas um códon cada.
Sobre ser universal, a ideia é que durante muito tempo se acreditou que o código genético era igual para todos os seres vivos. Porém, atualmente se reconhece que podem existir pequenas diferenças entre alguns organismos e também entre genes nucleares e mitocondriais.
A ideia principal é: o código genético é formado por 64 códons; 3 param a tradução e 61 codificam aminoácidos. Como há mais códons do que aminoácidos, vários códons podem indicar o mesmo aminoácido.
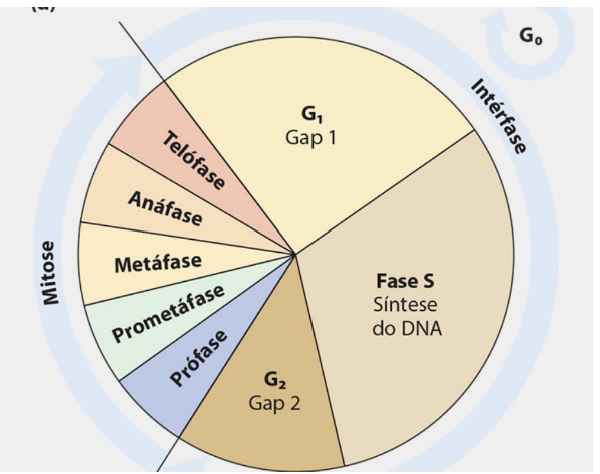
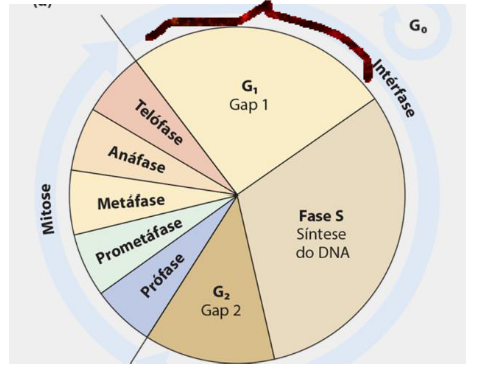
CICLO CELULAR
-
O crescimento e o desenvolvimento de cada organismo dependem, em grande parte da multiplicação, do crescimento e da diferenciação de suas células-filhas.
-
Todas as células tem capacidade de crescer e se reproduzir.
-
Nas células eucariontes isso ocorre de maneira cíclica, em um processo chamado ciclo celular.
-
O ciclo pode ser dividido em 2 grandes períodos.
-
Intérfase – A célula cresce, com aumento quantitativo de suas moléculas e material genético, e se prepara para a próxima divisão.
-
Mitose – A célula se divide (núcleo e citoplasma) em duas células-filhas.
-
No adulto, o processo de proliferação celular é importante para a reposição de células mortas e para a regeneração dos tecidos.
-
Ao mesmo tempo, ocorre um processo fisiológico de morte celular programada, denominado apoptose, que elimina células danificadas ou indesejáveis e mantem o número de células nos tecidos constante.
-
A intérfase corresponde a cerca de 95% do tempo total do ciclo celular, que dura geralmente de 12 a 24 horas.
-
Esse processo pode ser subdividido nas fases G1, S e G2.
O crescimento e o desenvolvimento de um organismo dependem muito do que acontece com suas células. Para o corpo crescer, renovar tecidos e manter suas funções, as células precisam crescer, se multiplicar e se diferenciar.
Nas células eucariontes, esse processo acontece de forma organizada e repetitiva, chamado de ciclo celular. O ciclo celular é como uma sequência de etapas que a célula passa até conseguir se dividir e formar novas células-filhas.
O ciclo celular pode ser dividido em dois grandes períodos: intérfase e mitose. A intérfase é a fase em que a célula cresce, aumenta suas moléculas, duplica seu material genético e se prepara para a divisão. Já a mitose é o momento em que a célula realmente se divide, formando duas células-filhas.
No adulto, a proliferação celular continua sendo importante. Ela serve para repor células mortas e ajudar na regeneração dos tecidos, como acontece, por exemplo, quando há renovação natural de células ou recuperação de uma lesão.
Ao mesmo tempo, o organismo também realiza a apoptose, que é uma morte celular programada. Isso significa que algumas células danificadas, velhas ou desnecessárias são eliminadas de forma controlada. Esse equilíbrio entre proliferação e apoptose ajuda a manter o número adequado de células nos tecidos.
A intérfase ocupa a maior parte do ciclo celular, cerca de 95% do tempo total. O ciclo completo geralmente dura de 12 a 24 horas, dependendo do tipo celular.
A intérfase é subdividida em três fases: G1, S e G2. De forma geral, na G1 a célula cresce, na S ocorre a duplicação do DNA, e na G2 a célula se prepara para entrar em divisão.
A ideia principal é: o ciclo celular organiza a vida da célula em preparação e divisão; primeiro vem a intérfase, com crescimento e duplicação do DNA, depois vem a mitose, formando duas células-filhas.
CLASSIFICAÇÃO DAS CÉLULAS ANIMAIS QUANTO A PROLIFERAÇÃO
-
Células de divisão contínua – Ex.: Células da medula óssea, do sistema linfático, do epitélio intestinal e dos folículos capilares.
-
Células que se dividem apenas em resposta a estímulos – Nesse caso, as células podem permanecer em um período chamado G0, que corresponde a um estado de dormência, na qual a célula mantem um baixo metabolismo e não se prolifera.
-
No entanto, se estimulada por fatores específicos, ela pode reingressar no ciclo celular na fase G1.
-
Ex.: Fibroblastos da pele, hepatocitos, células do pâncreas, do pulmão, do ovário, do rim, entre outros.
-
Células terminalmente diferenciados – neurônios e células musculares esqueléticas e cardíacas.
INTÉRFASE
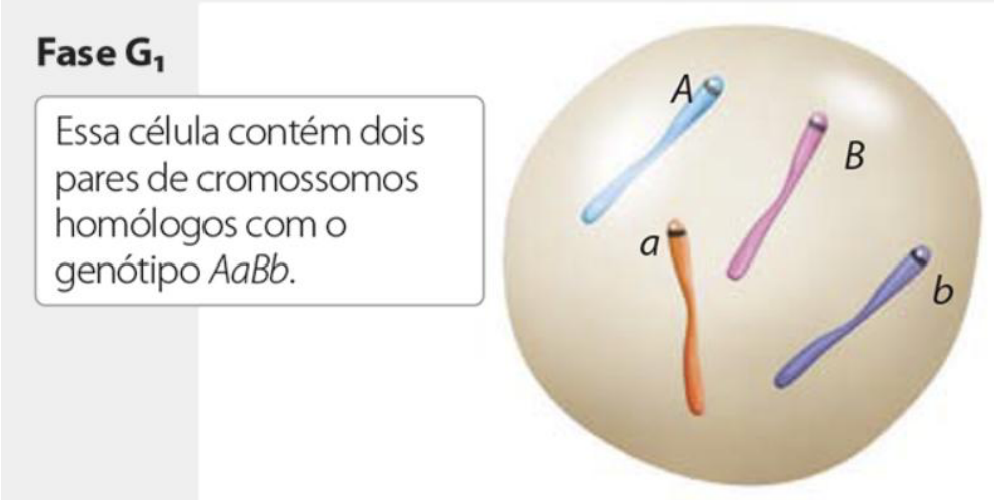
-
Período G1 – Nessa fase, inicia-se novamente a síntese de RNA (principalmente rRNA) e proteínas, que havia sido descontinuada durante a mitose.
-
Esse período é caracterizado pelo crescimento contínuo da célula.
-
Nessa fase, ocorre uma importante decisão: se a célula vai continuar se proliferando ou irá entrar em um estado quiescente (G0).
-
Os fatores de crescimento extracelulares vão sinalizar respostas intracelulares para influenciar nessa decisão.
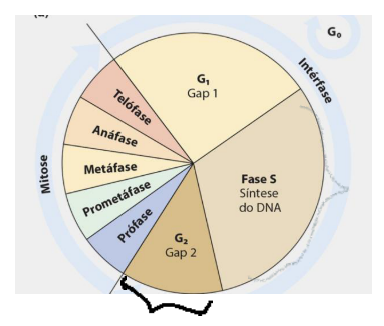
-
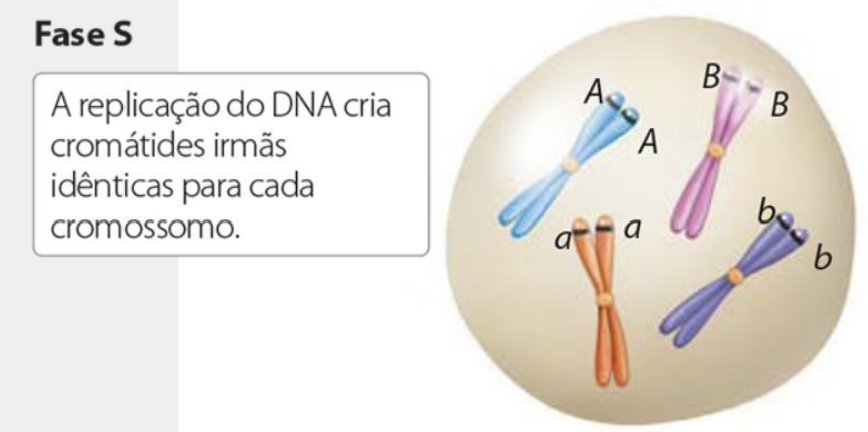
Período S – Replicação do DNA de todo o genoma da célula e duplicação da quantidade de histonas para que toda a cromatina seja duplicada.
-
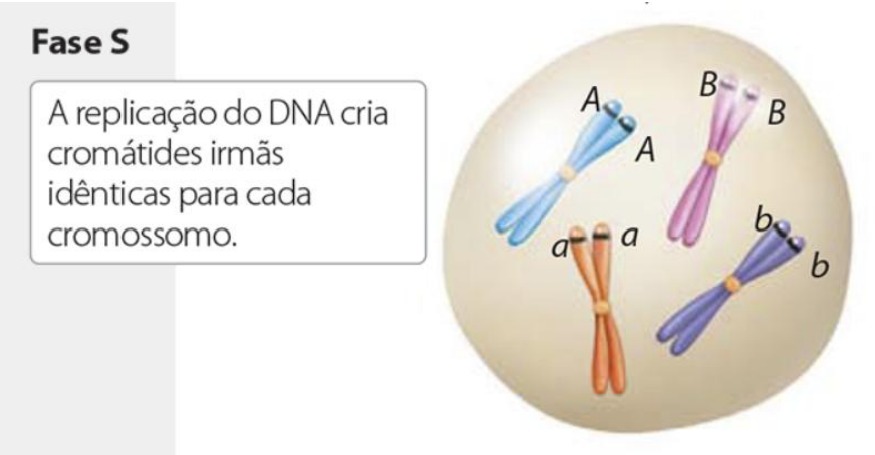
A replicação do DNA cria cromátides irmãs idênticas para cada cromossomo.
-
Período G2 – É uma fase crítica de checagem que antecede a mitose.
-
A célula permanece em um ponto de checagem em G2, até que a replicação tenha sido completada, e que possíveis erros do DNA tenham sido reparados antes que eles se transmitam para as células-filhas.
-
Também ocorre a síntese de proteínas, que serão necessárias para a condensação cromossômica durante a mitose.
A intérfase é o período em que a célula ainda não está se dividindo, mas está se preparando para isso. Ela é dividida em três fases principais: G1, S e G2.
No período G1, a célula retoma a produção de RNA e proteínas, principalmente porque essa atividade havia diminuído durante a mitose. Essa fase é marcada pelo crescimento da célula, ou seja, ela aumenta sua atividade e se prepara para continuar o ciclo.
Ainda em G1, acontece uma decisão importante: a célula pode continuar no ciclo celular e se preparar para dividir, ou pode entrar em G0, que é um estado de repouso, em que ela não se prolifera. Essa decisão depende de sinais, como os fatores de crescimento extracelulares, que influenciam respostas dentro da célula.
No período S, ocorre a replicação do DNA. Isso significa que todo o material genético da célula é duplicado. Além disso, também ocorre a duplicação da quantidade de histonas, porque o DNA duplicado precisa continuar organizado como cromatina.
A replicação do DNA forma as cromátides irmãs. Elas são cópias idênticas de cada cromossomo e serão separadas depois, durante a mitose, para que cada célula-filha receba o material genético correto.
No período G2, a célula faz uma checagem antes de entrar em mitose. Ela verifica se a replicação do DNA foi completada e se existem erros que precisam ser corrigidos antes da divisão.
Também em G2, ocorre a produção de proteínas importantes para a condensação cromossômica, que acontecerá na mitose. Ou seja, a célula prepara os cromossomos para ficarem mais compactados e organizados.
A ideia principal é: G1 é crescimento e decisão; S é duplicação do DNA; G2 é checagem e preparação final para a mitose.
MITOSE
-
A mitose consiste na divisão da célula em duas células-filhas.
-
Esse processo pode ser subdividido em 4 fases: Prófase, Metáfase, Anáfase e Telófase.
A mitose é o processo em que uma célula se divide e forma duas células-filhas. Essas células-filhas recebem o mesmo material genético da célula original.
Antes da mitose acontecer, o DNA já foi duplicado na intérfase, principalmente na fase S. Então, quando a célula entra em mitose, ela precisa apenas organizar e separar corretamente esse material duplicado.
ETAPAS DA MITOSE: PRÓFASE
-
Condensação das fibras de cromatina.
-
Dois pares de centríolo para os polos opostos da célula, para organizar os microtúbulos citoplasmáticos.
-
Interrupção da transcrição e uma desorganização dos nucléolos.
-
Com a ruptura do envoltório nuclear, os microtúbulos do fuso têm acesso aos cromossomos.
-
No citoplasma, os chamados centrossomos atuam formando feixes de microtúbulos, que vão constituir as fibras do fuso mitótico.

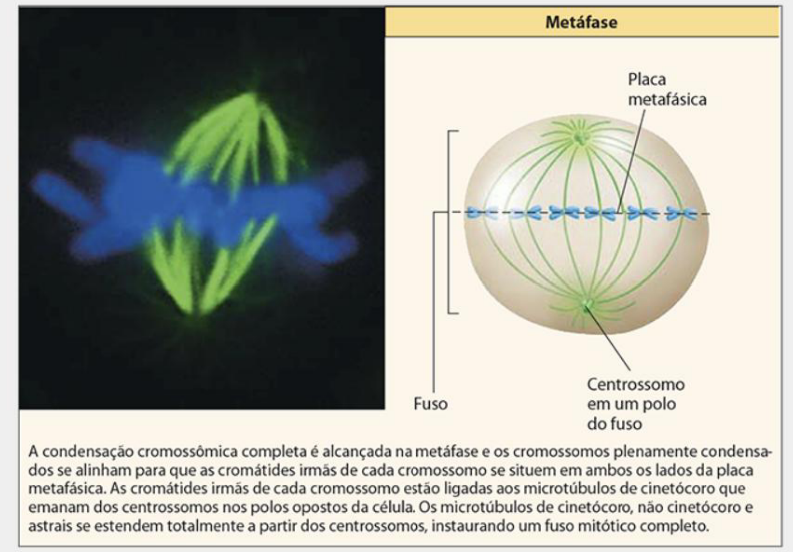
ETAPAS DA MITOSE: METÁFASE
-
Nesse estágio, a cromatina atinge seu grau máximo de condensação.
-
Os cromossomos são direcionados para a região equatorial da célula, formanda a placa metafásica.
-
O fuso é estabelecido entre os dois polos opostos da célula e a região equatorial.
-
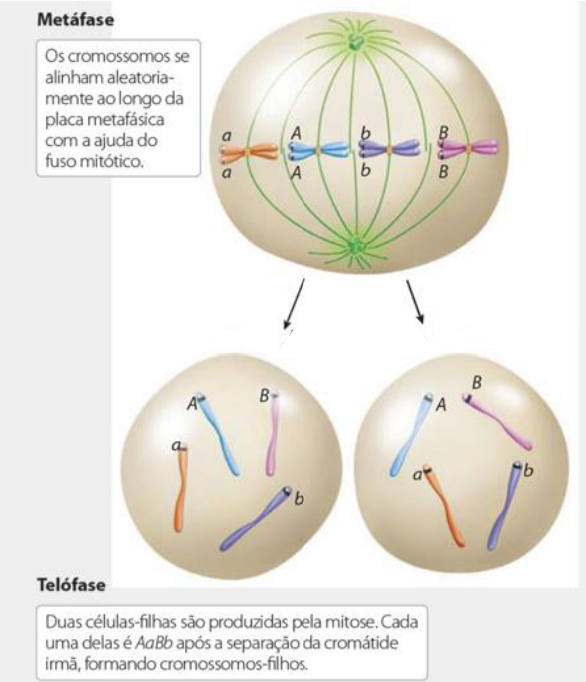
Os cromossomos se alinham aleatoriamente ao longo da placa metafásica com a ajuda do fuso mitótico.
-
A condensação cromossômica completa é alcançada na metáfase e os cromossomos plenamente condensados se alinham para que as cromátides irmãs de cada cromossomo se situem em ambos os lados da placa metafásica.
-
As cromátides irmãs de cada cromossomo estão ligadas aos microtúbulos de cinetócoro que emanam dos centrossomos nos polos opostos da célula.
-
Os microtúbulos de cinetócoro, não cinetócoro e astrais se estendem totalmente a partir dos centrossomos, instaurando um fuso mitótico completo.
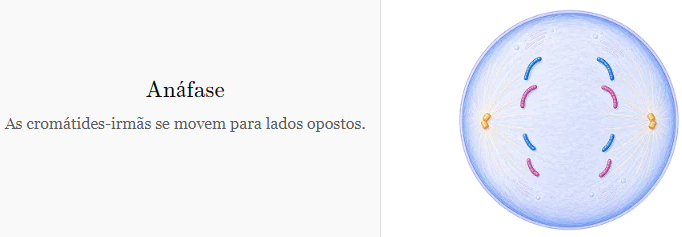
ETAPAS DA MITOSE: ANÁFASE
-
Fase caracterizada pela separação das cromátides-irmãs com a migração de cada uma delas para polos opostos.
-
As cromátides-irmãs, então, são chamadas de cromossomos-filhos.
-
Nessa migração, as fibras do fuso mitótico se encurtam, tracionando os cromossomos-filhos para os polos.
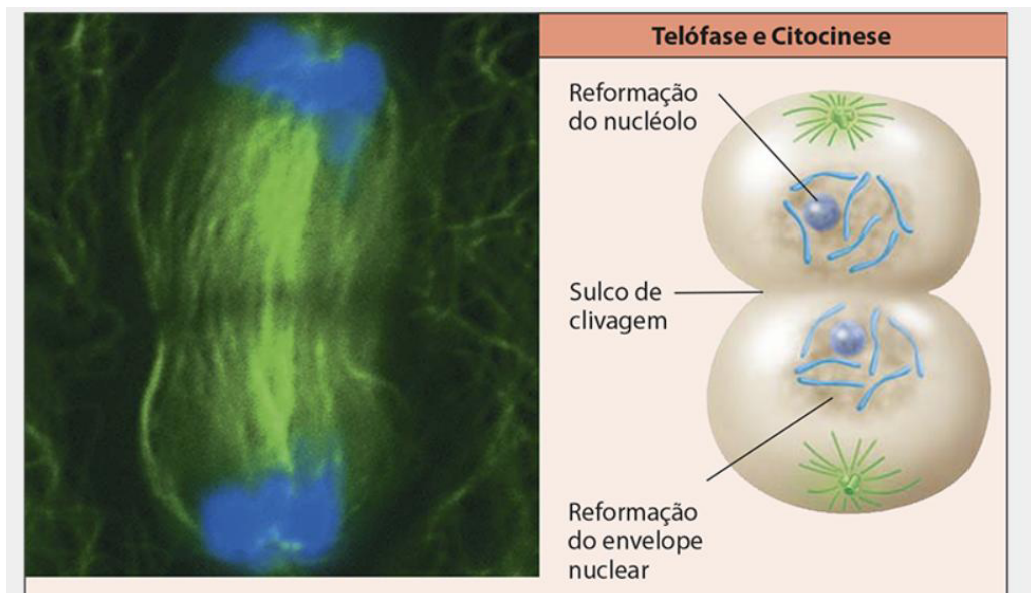
ETAPAS DA MITOSE: TELÓFASE E CITOCINESE
-
Quando os cromossomos-filhos atingem seus respectivos polos, ocorre o desaparecimento dos microtúbulos do fuso, tem início a Telófase.
-
Essa fase é caracterizada pela reconstituição do envoltório nuclear e descondensação da cromatina, formando os núcleos das células-filhas.
-
Além disso, ocorre a divisão citoplasmática, para que as duas células-filhas recebam partes iguais do conteúdo citoplasmático.
-
Ocorre a reformação do nucléolo e do envelope nuclear, assim como a formação de um sulco de clivagem.
-
Duas células-filhas são produzidas pela mitose, completando-se com a separação da cromátide irmã e formando cromossomos-filhos.

CHECKPOINTS
PONTO DE RESTRIÇÃO
-
Referem-se a pontos de parada durante o ciclo celular onde a estrutura do material genético é analisada.
-
Havendo algum erro e se ele for detectado, a célula entra em apoptose (morte programada).
-
Ocorre durante a fase G1.
Checkpoint G1
-
A proteína supressora tumoral p16 suprime a atividade do complexo ciclina D Cdk4 e há checagem da estrutura do DNA.
-
Caso a célula passe pelo checkpoint, o complexo ciclina-quinase torna-se ativo novamente.
Checkpoint G2
-
Esta pausa pré-mitótica imposta em G2 consiste em uma variedade de mecanismos de segurança que agem para controlar possíveis erros de replicação.
Checkpoint do fuso mitótico
-
Ocorre durante a metáfase, quando todos os cromossomos estão alinhados na placa equatorial.
-
Nesta etapa, ocorre o monitoramento da ligação dos cromossomos aos microtúbulos do fuso mitótico, visando impedir erros na segregação e garantir a integridade genômica a nível cromossômico.
Os checkpoints são pontos de controle do ciclo celular. Eles funcionam como “paradas de segurança”, em que a célula verifica se o material genético está adequado antes de continuar para a próxima etapa.
O ponto de restrição ocorre durante a fase G1. Nessa fase, a célula decide se vai continuar no ciclo celular, se dividir futuramente, ou se vai parar. É um momento importante porque a célula ainda pode impedir que um erro seja passado adiante.
Se algum erro importante no DNA for detectado e não puder ser corrigido, a célula pode entrar em apoptose, que é a morte celular programada. Isso é uma forma de proteção do organismo, porque impede que células com defeitos continuem se multiplicando.
No checkpoint G1, ocorre a checagem da estrutura do DNA. O material cita a proteína supressora tumoral p16, que reduz a atividade do complexo ciclina D–Cdk4. Em termos simples, isso ajuda a frear o ciclo celular enquanto a célula verifica se está tudo certo.
Se a célula passa pelo checkpoint, o complexo ciclina-quinase volta a ficar ativo, permitindo que o ciclo celular continue. Ou seja, se não houver problema grave, a célula segue para as próximas fases.
O checkpoint G2 acontece antes da mitose. Ele verifica se a replicação do DNA foi completada corretamente e se existem erros que precisam ser reparados antes da célula entrar em divisão.
Esse ponto é importante porque, se a célula entrasse em mitose com o DNA mal duplicado ou com erros, esses problemas poderiam ser transmitidos para as células-filhas.
O checkpoint do fuso mitótico ocorre durante a metáfase, quando os cromossomos estão alinhados na placa equatorial. Nessa fase, a célula verifica se os cromossomos estão corretamente ligados aos microtúbulos do fuso mitótico.
Esse controle evita erros na separação dos cromossomos durante a anáfase. Assim, cada célula-filha recebe a quantidade correta de material genético.
A ideia principal é: os checkpoints são mecanismos de segurança do ciclo celular; eles verificam o DNA e a ligação dos cromossomos ao fuso, impedindo que células com erros continuem se dividindo.
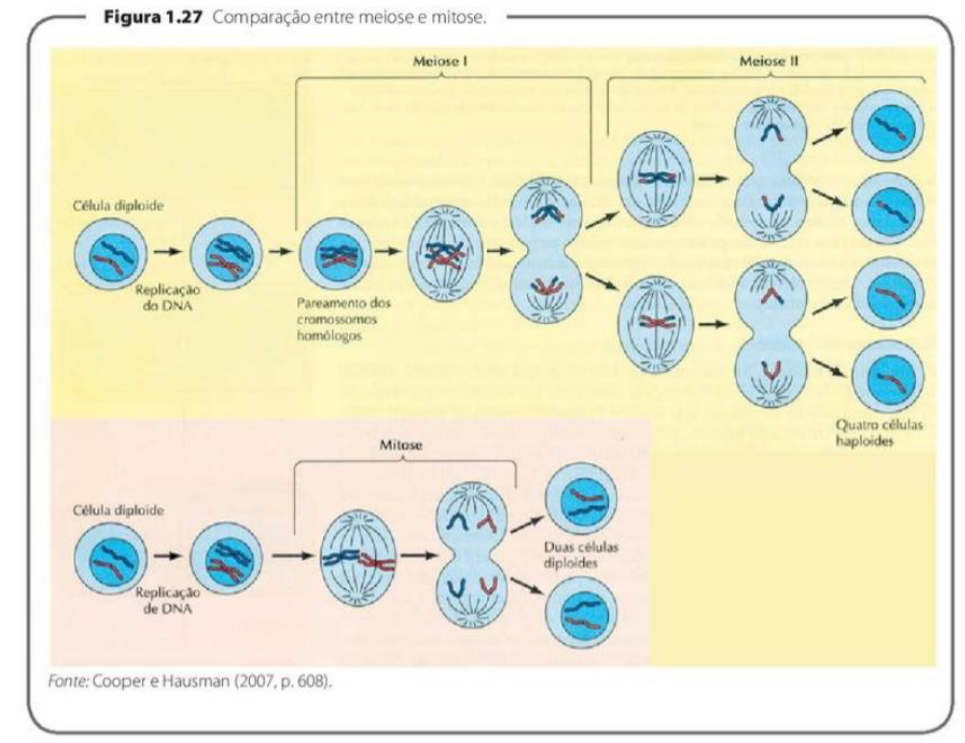
MEIOSE
-
A meiose ocorre nas células germinativas, permitindo a formação de células gaméticas que possuem somente a metade do número de cromossomos (são células haploides).
-
Na fecundação, a fusão de duas células gaméticas, forma, novamente, uma célula diploide para a origem de um novo organismo.
-
O processo de meiose compreende duas divisões: a meiose I (disjunção dos cromossomos homólogos) e a meiose II (disjunção das cromátides-irmãs).
-
Cada divisão é subdividida em: prófase, metáfase, anáfase e telófase.
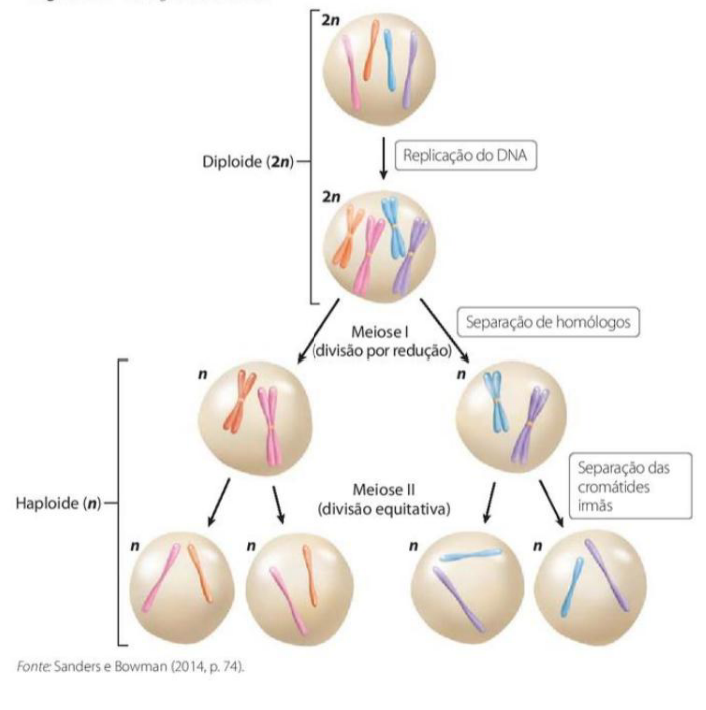
A meiose é um tipo de divisão celular que ocorre nas células germinativas, com o objetivo de formar os gametas, como óvulos e espermatozoides.
A principal característica da meiose é que ela reduz o número de cromossomos pela metade. Por isso, os gametas são chamados de células haploides, porque possuem apenas metade do número de cromossomos da célula original.
Na fecundação, acontece a união de dois gametas: um masculino e um feminino. Como cada um tem metade dos cromossomos, a fusão dos dois forma novamente uma célula diploide, com o número completo de cromossomos. Essa célula poderá dar origem a um novo organismo.
A meiose acontece em duas divisões seguidas: meiose I e meiose II.
Na meiose I, ocorre a separação dos cromossomos homólogos. Esses cromossomos formam pares, sendo um de origem materna e outro de origem paterna.
Na meiose II, ocorre a separação das cromátides-irmãs, de forma parecida com o que acontece na mitose.
Cada uma dessas divisões possui as fases: prófase, metáfase, anáfase e telófase. Por isso, temos prófase I, metáfase I, anáfase I e telófase I; depois prófase II, metáfase II, anáfase II e telófase II.
A ideia principal é: a meiose forma gametas haploides, com metade dos cromossomos, por meio de duas divisões celulares: na primeira separam-se os cromossomos homólogos; na segunda, as cromátides-irmãs.
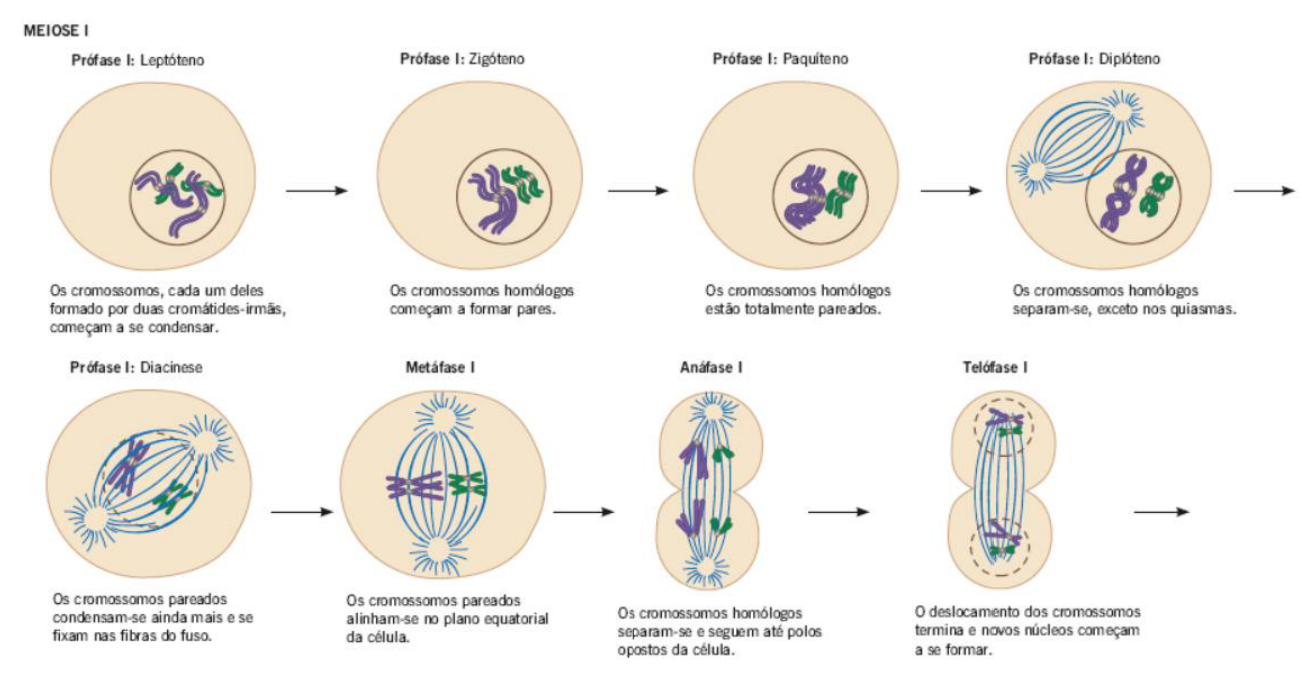
MEIOSE I – PRÓFASE I
-
A prófase I também inicia após o período S, em que o teor de DNA é duplicado de 2C para 4C.
-
Obs.: toda célula eucarionte inicia seu ciclo em G1 com uma quantidade de DNA igual a 2C.
-
Na prófase I, ocorre o pareamento dos cromossomos homólogos que não ocorre na mitose.
-
Cada núcleo filho, ao final da meiose I, irá receber um membro de cada par de cromossomos homólogos.
-
A prófase I é subdividida em 5 etapas: leptóteno, zigóteno, paquíteno, diplóteno e diacinese.
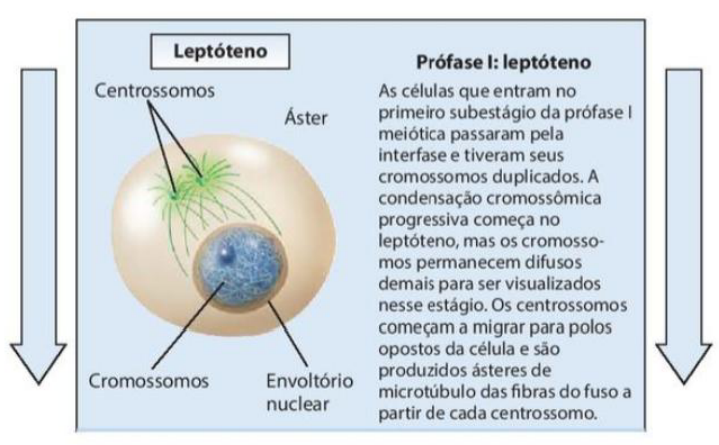
1. Leptóteno
-
As células que entram no primeiro subestágio da prófase I meiótica passaram pela interfase e tiveram seus cromossomos duplicados.
-
A condensação cromossômica progressiva começa no leptóteno, mas os cromossomos permanecem difusos demais para ser visualizados nesse estágio.
-
Os centrossomos começam a migrar para polos opostos da célula e são produzidos ásteres de microtúbulo das fibras do fuso a partir de cada centrossomo.
-
No leptóteno, ocorre a condensação gradual da cromatina em cromossomos.
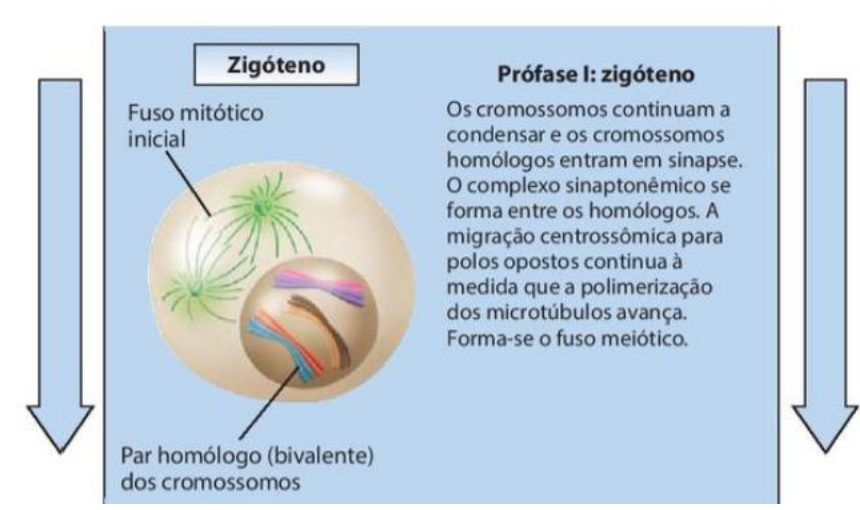
2. Zigóteno
-
Os cromossomos continuam a condensar e os cromossomos homólogos entram em sinapse.
-
O complexo sinaptonêmico se forma entre os homólogos.
-
A migração centrossômica para polos opostos continua à medida que a polimerização dos microtúbulos avança.
-
Forma-se o fuso meiótico.
-
No zigóteno, há a aproximação e o pareamento entre os cromossomos homólogos, em um processo chamado sinapse.
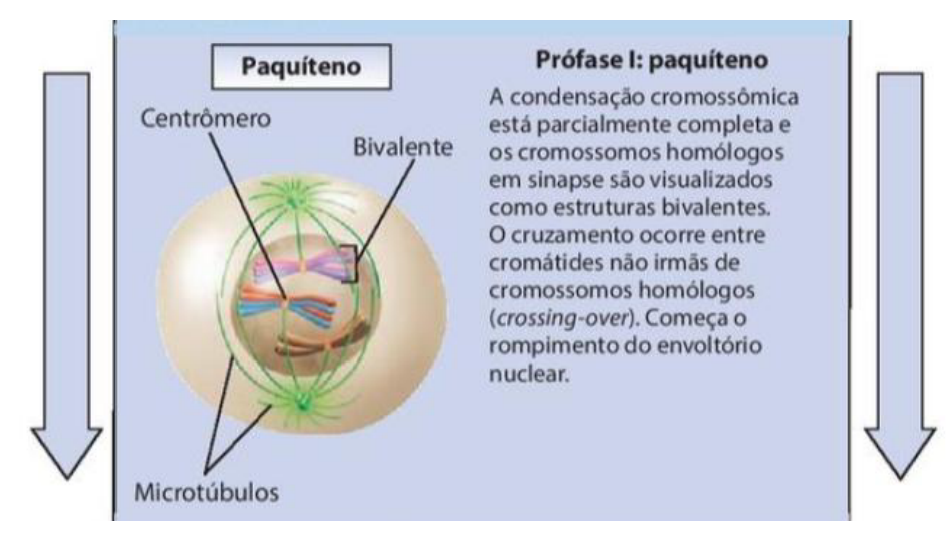
3. Paquíteno
-
A condensação cromossômica está parcialmente completa e os cromossomos homólogos em sinapse são visualizados como estruturas bivalentes.
-
O cruzamento ocorre entre cromátides não irmãs de cromossomos homólogos (crossing-over).
-
Começa o rompimento do envoltório nuclear.
-
No paquíteno, os cromossomos permanecem emparelhados e pode ocorre a troca de segmentos de DNA entre eles, na chamada recombinação genética, permuta ou crossing-over.
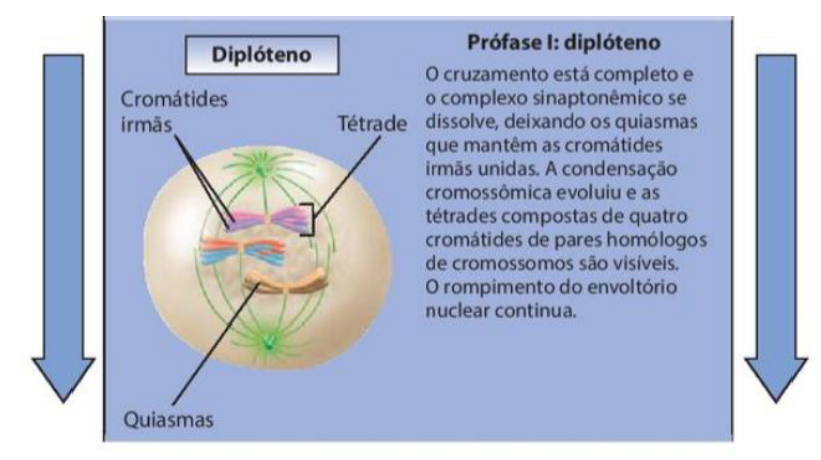
4. Diplóteno
-
O cruzamento está completo e o complexo sinaptonêmico se dissolve, deixando os quiasmas que mantêm as cromátides irmãs unidas.
-
A condensação cromossômica evoluiu e as tétrades compostas de quatro cromátides de pares homólogos de cromossomos são visíveis.
-
O rompimento do envoltório nuclear continua.
-
No diplóteno, a separação entre os cromossomos homólogos se inicia, mas não se completa.
-
Os cromossomos homólogos continuam ligados em alguns fragmentos denominados quiasmas (em forma de cruz).
-
Esses locais correspondem aos fragmentos que sofreram troca de DNA durante o paquíteno.
5. Diacinese
-
O fuso meiótico está bem estabelecido, com feixes de microtúbulos de cinetócoro amarrando os cromossomos homólogos das tétrades nos polos opostos.
-
O envelope nuclear está totalmente degradado.
-
As tétrades são movidas para o meio da célula.
-
Na diacinese, ocorre um aumento da separação entre os cromossomos, de forma que os quiasmas são deslocados para as extremidades.
-
Nessa etapa também ocorre a ruptura do envoltório nuclear e a ligação entre as fibras do fuso e os cromossomos homólogos, preparando para o início da metáfase.
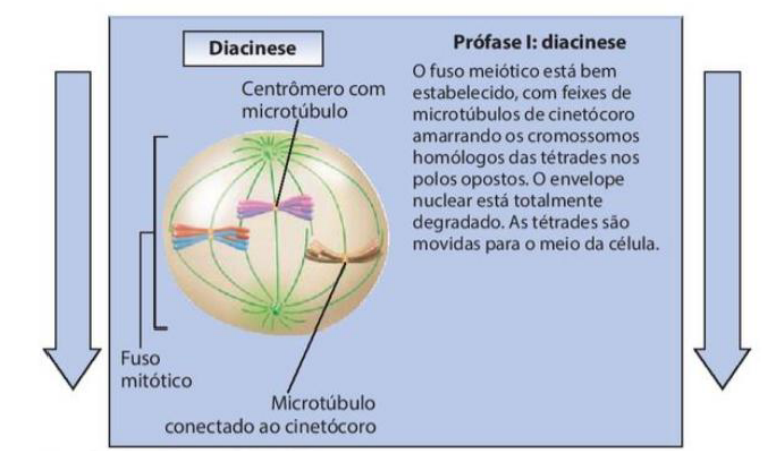
A prófase I é a primeira fase da meiose I e acontece depois da fase S da intérfase, quando o DNA já foi duplicado. Por isso, a célula passa de 2C para 4C, ou seja, ela continua com o mesmo número de cromossomos, mas agora com o DNA duplicado.
A grande diferença da prófase I da meiose em relação à mitose é que aqui ocorre o pareamento dos cromossomos homólogos. Esses cromossomos homólogos são os pares correspondentes, um de origem materna e outro de origem paterna. Esse pareamento não acontece na mitose.
Ao final da meiose I, cada célula-filha receberá apenas um cromossomo de cada par homólogo. Por isso, a meiose I é chamada de divisão reducional, pois reduz pela metade o número de cromossomos.
A prófase I é uma fase longa e importante, dividida em cinco etapas: leptóteno, zigóteno, paquíteno, diplóteno e diacinese.
No leptóteno, começa a condensação gradual da cromatina em cromossomos. Os cromossomos já foram duplicados, mas ainda estão finos e pouco visíveis. Também começa a movimentação dos centrossomos para lados opostos da célula.
No zigóteno, os cromossomos homólogos começam a se aproximar e se parear. Esse pareamento recebe o nome de sinapse. Nessa fase também se forma o complexo sinaptonêmico, que ajuda a manter os cromossomos homólogos unidos.
No paquíteno, os cromossomos homólogos já estão bem pareados e formam estruturas chamadas bivalentes. É aqui que pode ocorrer o crossing-over, também chamado de permuta ou recombinação genética. Nesse processo, cromátides não irmãs trocam pedaços de DNA, aumentando a variabilidade genética.
No diplóteno, os cromossomos homólogos começam a se separar, mas ainda permanecem ligados em alguns pontos chamados quiasmas. Esses quiasmas correspondem às regiões onde ocorreu o crossing-over no paquíteno.
Na diacinese, os cromossomos ficam ainda mais condensados e os quiasmas se deslocam para as extremidades. O envoltório nuclear se rompe, o fuso meiótico já está formado e as fibras do fuso se ligam aos cromossomos, preparando a célula para a metáfase I.
A ideia principal é: na prófase I ocorre o pareamento dos cromossomos homólogos e o crossing-over, que são eventos fundamentais da meiose e não acontecem na mitose.
Para memorizar a sequência:
Leptóteno: cromossomos começam a condensar.
Zigóteno: homólogos se pareiam.
Paquíteno: ocorre crossing-over.
Diplóteno: homólogos começam a se separar, mas ficam presos nos quiasmas.
Diacinese: cromossomos se condensam mais, núcleo se rompe e o fuso se liga aos cromossomos.
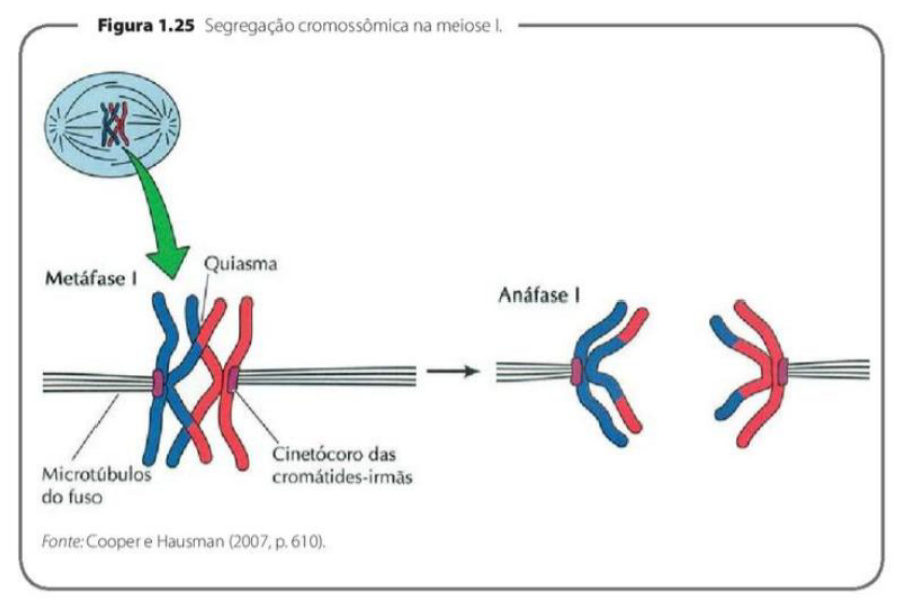
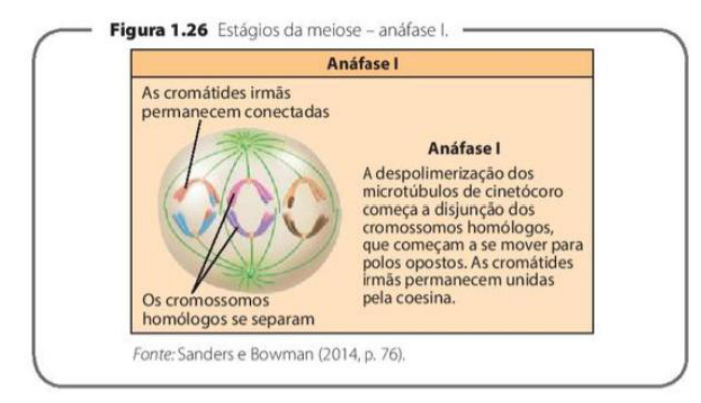
MEIOSE I – METÁFASE I, ANÁFASE I E TELÓFASE I
-
METÁFASE I: Os pares de cromossomos homólogos se dispõem na região equatorial das células, lembrando que cada cromossomo permanece com duas cromátides até o final da telófase I. Cada cromossomo do par se liga aos polos opostos da célula.
-
ANÁFASE I: Ocorre a movimentação de cada cromossomo do par de homólogos, de forma que em cada par os cromossomos materno e paterno (geralmente com segmentos trocados) se dirijam para polos diferentes. As cromátides-irmãs de cada cromossomo permanecem unidas pelo centrômero e migram juntas para um mesmo polo. A despolimerização dos microtúbulos de cinetócoro começa a disjunção dos cromossomos homólogos, que começam a se mover para polos opostos.
-
TELÓFASE I: Os grupos cromossômicos haploides chegam a seus respectivos polos, e logo é reconstruído o envoltório nuclear. O deslocamento dos cromossomos termina e novos núcleos começam a se formar. Em seguida, cada célula sofrerá a segunda divisão meiótica.
Na metáfase I, os pares de cromossomos homólogos ficam alinhados na região central da célula, chamada de região equatorial. Aqui é importante lembrar que cada cromossomo ainda está duplicado, ou seja, ainda possui duas cromátides-irmãs.
Nessa fase, cada cromossomo do par homólogo se liga a polos opostos da célula. Isso prepara a separação que acontecerá na próxima etapa. A diferença importante em relação à mitose é que, na metáfase I, quem se organiza no centro são os pares de homólogos, e não cromossomos individuais.
Na anáfase I, ocorre a separação dos cromossomos homólogos. Isso significa que o cromossomo de origem materna e o cromossomo de origem paterna vão para polos opostos da célula.
Mesmo com essa separação, as cromátides-irmãs permanecem unidas pelo centrômero. Então, na anáfase I, não se separam cromátides; separam-se cromossomos homólogos inteiros.
Esses cromossomos podem ter segmentos trocados por causa do crossing-over, que aconteceu antes, na prófase I. Por isso, eles podem não ser exatamente iguais aos cromossomos originais materno e paterno.
Na telófase I, os cromossomos chegam aos polos da célula. Cada polo recebe um conjunto haploide de cromossomos, ou seja, apenas um cromossomo de cada par homólogo.
Depois disso, o envoltório nuclear começa a se reorganizar e novos núcleos podem se formar. Em seguida, a célula se prepara para a meiose II, que será a segunda divisão meiótica.
A ideia principal é: na meiose I, os cromossomos homólogos se alinham, se separam e formam células com metade do número de cromossomos, mas cada cromossomo ainda continua com duas cromátides.
MEIOSE II
-
PRÓFASE II: Essa fase é de curta duração, mas é suficiente para ocorre novamente a montagem do fuso mitótico e o desaparecimento do envoltório nuclear.
-
METÁFASE II: Os cromossomos se dispõem na região equatorial e as cromátides-irmãs se prendem às fibras dos lados contrários do fuso.
-
ANÁFASE II: Nessa etapa ocorre a disjunção das cromátides-irmãs: elas são tracionadas para os polos opostos da célula.
-
TELÓFASE II: Divisão do citoplasma em duas células-filhas. As células-filhas haploides são separadas por membranas citoplasmáticas.
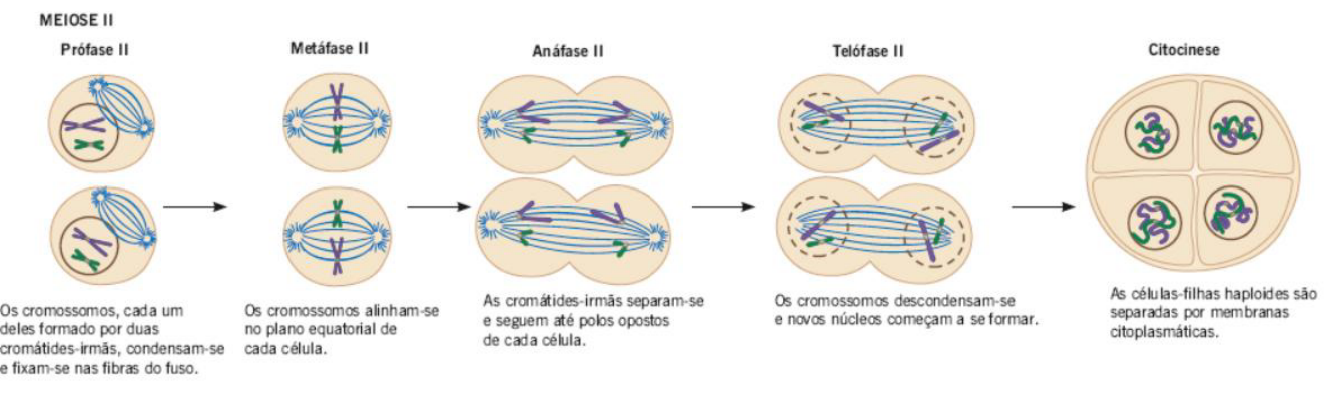
A meiose II é a segunda divisão da meiose. Ela acontece depois da meiose I e é parecida com a mitose, porque agora o que será separado são as cromátides-irmãs.
Na prófase II, ocorre novamente a organização do fuso e o desaparecimento do envoltório nuclear. Essa fase costuma ser curta, porque a célula já passou por uma grande preparação na meiose I.
Na metáfase II, os cromossomos se alinham na região equatorial da célula. Cada cromossomo ainda possui duas cromátides-irmãs, e essas cromátides ficam ligadas às fibras do fuso vindas de lados opostos.
Na anáfase II, ocorre a separação das cromátides-irmãs. Elas se separam no centrômero e são puxadas para polos opostos da célula.
Na telófase II, os cromossomos chegam aos polos e ocorre a divisão do citoplasma. Ao final, formam-se células-filhas haploides, ou seja, com metade do número de cromossomos.
A ideia principal é: na meiose I separam-se os cromossomos homólogos; na meiose II separam-se as cromátides-irmãs.
LISTA DE EXERCÍCIOS QUE A PROF PASSOU, COM RESPOSTAS
1) DEFINA O QUE SÃO ÁCIDOS NUCLÉICOS E COMO SÃO CONSTITUÍDOS.
Resposta: Os ácidos nucleicos (DNA e RNA) são polímeros lineares longos que armazenam a informação genética e desempenham diversas funções cruciais no organismo. Eles consistem em um grande número de unidades básicas chamadas nucleotídeos, ligados uns aos outros. Cada nucleotídeo, por sua vez, é formado por três partes: uma pentose (um tipo de açúcar, como ribose ou desoxirribose), um grupamento fosfato e uma base nitrogenada.
2) O QUE É O PROCESSO DE REPLICAÇÃO E QUAIS ENZIMAS FAZEM PARTE DESSE PROCESSO? EXPLIQUE A FUNÇÃO DE CADA UMA.
Resposta: A replicação é o processo essencial para a duplicação do material genético, ocorrendo durante a divisão celular para garantir que o mesmo material original seja repassado às células-filhas. As principais enzimas envolvidas são:
-
Helicases: Realizam a abertura da dupla hélice do DNA, permitindo a movimentação da forquilha de replicação.
-
Primases: Sintetizam e inserem os primers (iniciadores), que são segmentos curtos de ácido nucleico associados à fita molde, fornecendo a extremidade necessária para iniciar a nova fita.
-
DNA Polimerases: Realizam a incorporação dos novos nucleotídeos sequencialmente na extremidade 3′-OH livre da cadeia em formação.
-
Topoisomerases: Reduzem a tensão gerada pelas supertorções causadas pela abertura da hélice, induzindo uma quebra transitória na cadeia.
-
SSB (Single-stranded DNA binding proteins): Proteínas que se ligam às fitas simples de DNA, mantendo-as separadas e impedindo que se unam novamente.
- DNA Ligase: Une os fragmentos de Okasaki gerados na fita descontínua, completando a fita.
3) O QUE É O PROCESSO DE TRANSCRIÇÃO? DETALHE COMO OCORRE A IDENTIFICAÇÃO DE ONDE ELA DEVE SER INICIADA E FINALIZADA.
Resposta: A transcrição é o processo pelo qual uma molécula de RNA é sintetizada utilizando uma das fitas da molécula de DNA como molde.
-
Início: A enzima RNA polimerase identifica o local onde deve iniciar a transcrição ao reconhecer uma região específica do DNA chamada promotor. Entre essas marcações, destaca-se a sequência TATA box, localizada antes dos genes, cuja associação com a enzima forma o complexo de pré-iniciação para a síntese do RNA.
-
Finalização: A RNA polimerase identifica o local onde deve parar de sintetizar o RNA ao encontrar sequências genéticas específicas denominadas finalizadores. Após essa leitura, ocorre a dissociação da RNA polimerase do DNA, liberando a nova molécula de RNA recém-transcrita.
4) O QUE É TRADUÇÃO E QUAL O PAPEL DOS RNAS NESTE PROCESSO?
Resposta: A tradução é a fase em que a informação contida no RNA é “lida” pelos ribossomos para a montagem de uma proteína (cadeia polipeptídica) por meio da ligação de aminoácidos.
-
RNA mensageiro (mRNA): Leva a informação genética (códons) copiada do DNA até os ribossomos e é a única molécula efetivamente “traduzida” em proteína.
-
RNA ribossômico (rRNA): Constitui o principal componente estrutural dos ribossomos, sendo rico em pareamentos internos e fundamental para acoplar os demais RNAs no sítio de tradução.
-
RNA transportador (tRNA): Capta os aminoácidos dispersos na célula e os transporta fisicamente até o ribossomo. Ele possui o anticódon, que emparelha com o códon do mRNA e garante que o aminoácido correto seja inserido na proteína.
5) A TABELA ABAIXO DEMONSTRA A SEQUÊNCIA DE AMINOÁCIDOS DE UM SEGMENTO DE UMA PROTEÍNA E OS RESPECTIVOS ANTICÓDONS DOS RNA TRANSPORTADORES (ADAPTADO: MUNDO EDUCAÇÃO, 2026): […] QUAL A SEQUÊNCIA DE CÓDONS DO RNA MENSAGEIRO QUE PARTICIPOU DESSA TRADUÇÃO?
Resposta: Sabendo que o anticódon do tRNA é complementar à sequência do códon no mRNA e que no RNA a base Adenina (A) pareia com Uracila (U), e a Citosina (C) pareia com Guanina (G), chegamos à seguinte dedução:
-
Anticódon AAG -> Códon UUC
-
Anticódon AAC -> Códon UUG
-
Anticódon AUG -> Códon UAC
-
Anticódon GGA -> Códon CCU
-
Anticódon GCG -> Códon CGC
Sequência de códons do mRNA: UUC – UUG – UAC – CCU – CGC.
6) ANALISE AS SEQUÊNCIAS A SEGUIR, AS QUAIS SE REFEREM A MOLÉCULAS DE RNA MENSAGEIROS OBTIDAS A PARTIR DE CÉLULAS PERTENCENTES A DOIS ORGANISMOS DIFERENTES (ADAPTADO: DESCOMPLICA, 2026): […] COM BASE NESTA INFORMAÇÃO, CONSULTE A TABELA DE CÓDIGO GENÉTICO E ESCREVA A CADEIA DE AMINOÁCIDOS OBTIDA DA TRADUÇÃO DAS MOLÉCULAS DE RNA MENSAGEIROS DOS ORGANISMOS 1 E 2.
Resposta: Ao consultar a tabela do código genético, traduzimos as sequências de três em três bases (códons):
-
Organismo 1 (CCU-GCU-GGC-ACA):
CCU = Prolina (Pro) | GCU = Alanina (Ala) | GGC = Glicina (Gly) | ACA = Treonina (Thr).
Cadeia do organismo 1: Pro – Ala – Gly – Thr
-
Organismo 2 (CCA-GCG-GGU-ACU):
CCA = Prolina (Pro) | GCG = Alanina (Ala) | GGU = Glicina (Gly) | ACU = Treonina (Thr).
Cadeia do organismo 2: Pro – Ala – Gly – Thr
Nota: O fato de as duas sequências genéticas diferentes produzirem a mesmíssima proteína ilustra que o código genético é redundante, isto é, um mesmo aminoácido pode ser codificado por diferentes trincas (códons).
7) O QUE É MITOSE E MEIOSE? DIFERENCIE ESSES PROCESSOS.
Resposta: * Mitose: É a divisão celular padrão onde a célula (núcleo e citoplasma) se divide em duas células-filhas diploides e geneticamente idênticas à original. É a base para a proliferação, reposição de células mortas e regeneração de tecidos do corpo.
-
Meiose: É um processo que ocorre estritamente nas células germinativas para formar as células gaméticas (haploides), que levam apenas a metade do número de cromossomos e farão parte da reprodução.
-
Diferenciação: A mitose envolve apenas uma etapa de divisão que mantém o número total de cromossomos (2n) sem troca de DNA entre eles, gerando 2 células. A meiose abrange duas divisões consecutivas, produzindo 4 células de número cromossômico reduzido (n). Diferente da mitose, na meiose ocorre o pareamento dos cromossomos homólogos e a recombinação genética por crossing-over.
8) QUAIS ETAPAS EXISTEM NA MITOSE E NA MEIOSE.
Resposta:
-
Etapas da Mitose: É dividida em 4 fases sequenciais principais: Prófase, Metáfase, Anáfase e Telófase.
-
Etapas da Meiose: Compreende duas grandes divisões. A Meiose I (fase reducional) engloba: Prófase I, Metáfase I, Anáfase I e Telófase I. Onde a Prófase I possui cinco subetapas importantes: Leptóteno, Zigóteno, Paquíteno, Diplóteno e Diacinese. A Meiose II (fase equitativa) possui: Prófase II, Metáfase II, Anáfase II e Telófase II.
9) O QUE É GAMETOGÊNESE?
Resposta: A gametogênese é o processo biológico no qual as células germinativas sofrem sucessivas divisões, culminando na meiose, para permitir a formação de células gaméticas. Essas células, sendo haploides (com metade do DNA), têm como fim a fusão na fecundação, para originar e reestabelecer o material genético diploide em um novo organismo.
10) QUAL A DIFERENÇA ENTRE GAMETOGÊNESE FEMININA E MASCULINA?
Resposta: Embora ambas possuam como finalidade a formação de gametas baseados no processo de meiose para redução do número cromossômico, existem variações biológicas profundas que não estão listadas nas notas de aula fornecidas. Geralmente:
-
Gametogênese Masculina (Espermatogênese): É um processo relativamente ininterrupto (a partir da puberdade) que gera quatro espermatozoides viáveis com distribuição equitativa de citoplasma para cada célula-mãe germinativa meiótica.
-
Gametogênese Feminina (Ovogênese): Inicia-se de forma prematura na vida embrionária e sofre longas pausas (a divisão celular trava em etapas específicas). A divisão do citoplasma é bastante assimétrica, com a maior parte da reserva biológica sendo dedicada à formação de um único óvulo viável, com o descarte dos corpúsculos polares inviáveis.
LINK QUIZZ DO GEMINI: https://gemini.google.com/share/f126f814cea1